
Natural Resources
Conservation Service
Ecological site R015XF005CA
Steep Loamy Foothills
Accessed: 04/03/2026
General information
Approved. An approved ecological site description has undergone quality control and quality assurance review. It contains a working state and transition model, enough information to identify the ecological site, and full documentation for all ecosystem states contained in the state and transition model.
Figure 1. Mapped extent
Areas shown in blue indicate the maximum mapped extent of this ecological site. Other ecological sites likely occur within the highlighted areas. It is also possible for this ecological site to occur outside of highlighted areas if detailed soil survey has not been completed or recently updated.
MLRA notes
Major Land Resource Area (MLRA): 015X–Central California Coast Range
Major Land Resource Area (MLRA) 15 (Central California Coast Range): The MLRA is an area of gently sloping to steep, low mountains. Precipitation is evenly distributed throughout fall, winter, and spring but is very low in summer. Elevation ranges from sea level to 2,650 feet (810 meters) in most of the area, but up to 4,950 feet (1,510 meters) in some of the mountains. The soils in the area dominantly have a thermic soil temperature regime, a xeric soil moisture regime, and mixed or smectitic mineralogy.
Land Resource Unit (LRU) 15XF: The LRU includes Blue Ridge in the northern California Coast Ranges and steep hills east of Blue Ridge and east of the Stony Creek fault, extending north to the Klamath Mountains down to the southern portion of Napa and Yolo Counties. The LRU is formed mostly from Jurassic and Cretaceous sandstone and shale facies of the Great Valley sequence. This area includes north to south trending foothill slopes and alluvial back valleys. Soil temperature regime is mostly thermic, with some high elevation areas that are mesic, and soil moisture regime is xeric. Common vegetation includes introduced annual grasses and forbs, blue oak, chamise, ceanothus, manzanita and foothill pine. Elevations generally range from 500 to 2400 feet. Rainfall levels drop quickly from the mountains to the foothills and valley due to the rain shadow effect. Annual precipitation generally averages from 16 to 40 inches. LRU 15XF has several Environmental Protection Agency (EPA) Ecoregions; this ecological site is located in Ecoregion “6f”.
Classification relationships
This blue oak dominated site may include the following Allen-Diaz Class: 1) Blue Oak-Foothill Pine/Grass (Allen-Diaz et al., 1989). This site includes Blue Oak Woodland (BOW) and Blue Oak-Foothill Pine (BOP) of the California Wildlife Habitat Relationships System. The Society for Range Management Cover Type for this site is Blue Oak Woodland (Shiflet (Ed.), 1994). This site includes the Quercus douglasii Woodland Alliance from the publication "A Manual of California Vegetation, 2nd Edition", (Sawyer et al., 2009).
Ecological site concept
This ecological site predominantly occurs on moderately deep loamy soils found on steep to very steep foothill shoulders and backslopes. Steep slopes incur more erosion as water drains rapidly from these sites. Soils are also drier due to increased water runoff from this site. Found primarily on east and northwest facing aspects, this site receives peak solar radiation during the morning and at mid-day (Kimmins, 1987), experiencing lower evapotranspiration rates than south facing slopes.
The diagnostic soil horizon includes the presence of an Argillic Horizon or a zone of clay accumulation that indicate site stability. Permeability is slow, runoff high and erosion hazard is high.
This ecological site is an oak savanna with an understory dominated by non-native annual grasses including wild oat (Avena fatua) or slender oat (Avena barbata), a low to moderate cover of blue oak (Quercus douglasii), and California foothill pine (Pinus sabiniana), and shrub cover of Pacific poison oak (Toxicodendron diversiloba).
Associated sites
| R015XF002CA |
Clayey Foothills The Clayey Foothills ecological site is often found in association with the Steep Loamy Foothill Ecological Site, occurring predominantly on adjacent steep backslopes and footslopes. Vegetation is Blue oak with whiteleaf manzanita in the understory, along with annual grasses and forbs. The Clayey Foothills ecological site is similar to this site but differs in that its soil texture is clayey instead of loamy, and deep cracks develop in the soil surface that extend well down into the soil profile. Steep Loamy Foothills is found at higher elevations and generally has a greater amount and diversity of shrubs than the Clayey Foothills ecological site. |
|---|---|
| R015XF004CA |
Shallow Loamy Foothills The Shallow Loamy Foothills ecological site is found in association with this site, primarily on south and southwest facing slopes. Vegetation is Blue oak with a shrub layer of buckbrush, and an understory dominated by wild oat. |
Table 1. Dominant plant species
| Tree |
(1) Quercus douglasii |
|---|---|
| Shrub |
(1) Toxicodendron diversilobum |
| Herbaceous |
(1) Avena fatua |
Physiographic features
This ecological site predominantly occurs on moderately deep soils found on steep to very steep foothill shoulders and backslopes (See the Physiographic Diagram below). This ecological site is often found in association with another site, R015XF004CA, Shallow Loamy Foothills. This ecological site occurs predominantly at elevations of 1,000 to 2,000 feet however elevations may range from 300 to 2,000 feet. This site occurs on all aspects however the Reference Community Phase is primarily found on east to northwest facing slopes.
Slopes generally range from 30 to 60 percent however the full slope range is from 10 to 75 percent.
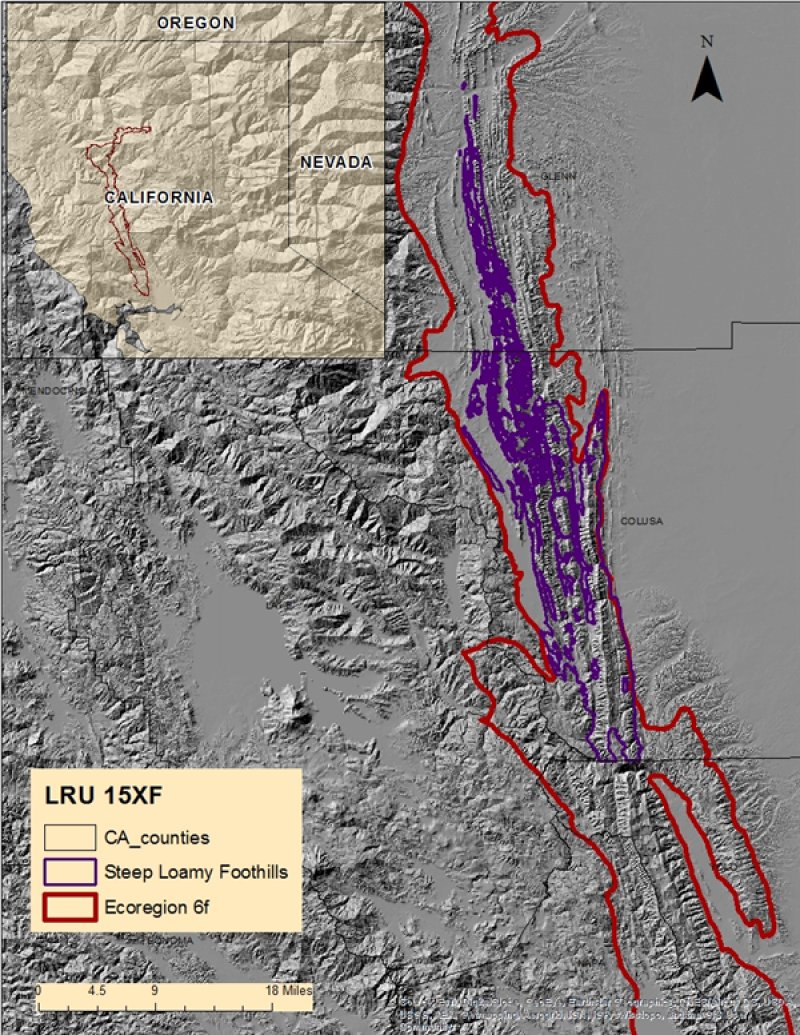
Figure 2. R015XF005CA - Steep Loamy Foothills
Table 2. Representative physiographic features
| Landforms |
(1)
Hill
|
|---|---|
| Flooding frequency | None |
| Ponding frequency | None |
| Elevation | 1,000 – 2,000 ft |
| Slope | 30 – 60% |
| Aspect | E, W, NW |
Climatic features
This ecological site has a Mediterranean climate characterized by hot summer temperatures and cool moist winters. The driest time of the year is generally in June, July, August and September. Precipitation falls primarily as rain during October through May. The northern part of the California Central Valley receives precipitation from winter storms from the Pacific Northwest. The timing, length and intensity of storms are highly variable and unpredictable.
Periodic drought may occur for months or years at a time, depending on the fluctuations of winds and ocean currents in the equatorial region of the Pacific ocean (Quinn and Keely, 2006).
The mean annual precipitation is 19 to 24 inches and the mean annual air temperature is 59 to 63 degrees Fahrenheit. The frost free period averages 224 days. The freeze free period averages 279 days.
Two climate stations were utilized due their central location within the ecological site: East Park Reservoir and Stony Gorge Reservoir. The southern extent of this site is subject to higher precipitation with increased elevation.
Table 3. Representative climatic features
| Frost-free period (average) | 224 days |
|---|---|
| Freeze-free period (average) | 279 days |
| Precipitation total (average) | 24 in |
Figure 3. Monthly precipitation range
Figure 4. Monthly average minimum and maximum temperature
Figure 5. Annual precipitation pattern
Figure 6. Annual average temperature pattern
Climate stations used
-
(1) STONY GORGE RSVR [USC00048587], Elk Creek, CA
-
(2) EAST PARK RSVR [USC00042640], Stonyford, CA
Influencing water features
The presence of an Argillic Horizon, steep slopes, and topographic position affect the rate of infiltration and runoff from this ecological site. The slowly permeable argillic layer affects water infiltration through subsurface layers and may alter interflow to downslope positions. Steep slopes and the site’s location on shoulders and backslope positions contribute to medium to very high runoff. The concave downslope and across slope shape also favors runoff from this site. The low surface litter layer and the amount of bare ground may also contribute to increases in overland flow and soil loss.
Soil features
The soils typically associated with this ecological site are formed in residuum from Cretaceous sandstone and shale facies of the Great Valley sequence. This soil series, Contra costa (Fine, mixed, superactive, thermic Mollic Haploxeralfs), has a moderate degree of soil development, and is moderately deep to hard bedrock.
Dominant soils are loamy throughout the soil profile. Surface textures are loam and clay loam and subsurface textures are clay, clay loam, channery clay loam and very gravelly clay loam. Available water capacity tends to be low to moderate on these sites (4-5 inches). These soils are well drained. Surface gravels range from 1 to 13 percent and subsurface gravels range from 1 to 21 percent. Subsurface fragments greater than 3 inches range from 0 to 6 percent.
The soils on this ecological site have an argillic horizon, a subsurface horizon of clay accumulation from illuviation. The presence of an argillic subsurface horizon indicates that the geomorphic surface has been relatively stable and that the period of stability is long (USDA, NRCS, 1999). Permeability is slow and runoff is medium to very high.
This ecological site is correlated with the following map units and components in MLRA 15:
Contra costa - Fine, mixed, superactive, thermic Mollic Haploxeralf
Soils Map Units:
CA011; Colusa County, California:
241; Contra costa–Altamont association, 30 to 50 percent slopes; Contra costa
305; Contra costa loam, 50 to 75 percent slopes
334; Millsholm-Contra costa association 30 to 75 percent slopes; Contra costa
CA21; Glenn County, California:
CtE; Contra costa clay loam 30 to 65 percent slopes
CvE; Contra costa–Millsholm clay loams, 30 to 65 percent, Contra costa

Figure 7. Contra costa Soil
Table 4. Representative soil features
| Parent material |
(1)
Residuum
–
sandstone and shale
|
|---|---|
| Surface texture |
(1) Loam (2) Clay loam |
| Family particle size |
(1) Clayey |
| Drainage class | Well drained |
| Permeability class | Slow |
| Soil depth | 20 – 40 in |
| Surface fragment cover <=3" | 1 – 13% |
| Surface fragment cover >3" | Not specified |
| Available water capacity (0-40in) |
4 – 5 in |
| Calcium carbonate equivalent (0-40in) |
1% |
| Electrical conductivity (0-40in) |
2 mmhos/cm |
| Sodium adsorption ratio (0-40in) |
2 |
| Soil reaction (1:1 water) (0-40in) |
5.6 – 7.3 |
| Subsurface fragment volume <=3" (Depth not specified) |
1 – 21% |
| Subsurface fragment volume >3" (Depth not specified) |
6% |
Ecological dynamics
Disturbance is defined as “any relatively discrete event in time that disrupts ecosystem, community, or population structure and changes resource pools, substrate availability, or the physical environment” (Pickett and White, 1985); a disturbance may be natural or anthropogenic in origin.
This ecological dynamics section will focus on the primary vegetation types found in this ecological site: Blue oak-California foothill pine and Blue oak Woodland. Significant areas of chamise and to a lesser extent Lower Montane Mixed Chaparral exist in the southern portion of the site. These areas are not considered part of this site, and therefore are not addressed within the ecological dynamics section
Historic Disturbances:
The interaction of several disturbance agents including fire, clearing, invasive species and drought has influenced and shaped the oak savanna environment. In the mid-1800s prior to European settlement fire frequency was approximately every 25 years (McClaran 1986). Native Americans regularly used fire to manage vegetation communities to provide food and fiber (Blackburn and Anderson 1993; McCleary 2004). The historic vegetation community likely experienced an understory fire regime (Arno, Allison-Burnell, 2002). Frequent low intensity fire likely left widely spaced overstory trees and removed smaller trees and brush (McCleary, 2004; Pavlik 1991; Mensing 1992; Stephens 1997). Following settlement before and after the gold rush, fires were more frequent, every 5 to 15 years, due to the intentional use of fire by ranchers and others to reduce brush. Shrub and oak recruitment increased in the absence of periodic fire or grazing in some environments (Purcell and Stephens, 2005).
Clearing of oaks occurred throughout the 1880’s for agriculture and livestock purposes (McCleary, 2004). Increased settlement also resulted in the loss of oaks in the support of fuels for railroads, mines and steamships. After the Second World War, there was extensive conversion of woodland to pasture, and the inventory of hardwood forest types in California has decreased with widespread conversion from residential and commercial development. Within the State of California about 1.9 million acres of hardwoods and chaparral were reported to have been cleared in rangeland improvement projects (Bolsinger, 1988). Chaining of oaks occurred during the 1960’s in an attempt to provide for more grassland production. The increase in grass production was short-lived (less than 10-20 years) following oak removal and oak representation on some landscapes was altogether eliminated. Foothill pine also suffered the same fate in some areas. Some research indicates that oak removal results in a rapid decline in soil quality, including a loss in soil organic matter and nitrogen (Dalgren et al., 2003), and that oaks help retain more water on site and enhance soil quality through nutrient cycling, organic matter deposition and reduced bulk density (O’Geen et al., 2010). Firewood cutting for fuel and residential and commercial development continue to contribute to the loss of oaks, though at a much slower rate. The lack of natural regeneration of oak has been attributed to many factors including herbivory, acorn predation, competition from annual grasses, and altered fire regimes (Fryer 2007; UC 2007; Sweitzer and Van Vuren 2002).
Drought: California grasslands experience an annual summer water deficit (Barbour and Major, 1977) as a result of the Mediterranean-influenced climate. This water deficit in combination with periodic drought can lead to changes in grassland species composition and production as a result of prolonged low water availability (Stromberg et al., 2007). The most recent drought period, now in its fourth year is unprecedented in California’s climate record (Griffin and Anchukaitis, 2014). Increased temperature and evaporation will likely have a significant effect on species composition and productivity on this site, favoring more droughty species, lowering oak seedling survival (Grünzweig et al., 2008) and lessening overall range production. Oaks are efficient water users; they are adapted to very low moisture conditions by virtue of their small leaf size, the regulation of water loss through the leaf stomata and by tapping into water below fractured rock (Baldocchi et al., 2007). Oak leaves may brown and drop prematurely during a drought year and fully recover the following year, potentially affecting the acorn crop, however, prolonged severe drought can result in mortality in oaks, especially on south facing slopes (Harper et al., 1991). Foothill pine has the ability to grow on the doughtiest sites by photosynthesizing during winter and spring and through low transpiration rates (Burns and Honkala (eds.), 1990).
Fire: Active fire suppression during the last century has allowed for the accumulation of fuels and a trend towards larger more devastating fires (McCleary 2004; Arno and Allison-Bunnell, 2002). The lack of fire on this site has resulted in significant regeneration and survival of California foothill pine, adding to ladder fuels and the increased fire risk. Foothill pine is susceptible to fire due to its thin flaky bark and heavy resin production (Howard, 1992). Blue oak can withstand low-intensity ground fires but is very susceptible to moderate to severe fire (Pavlik et al., 1992). Blue oak is considered a weak sprouter following fire, and sprouting declines with age (Burns and Honkala 1990, McDonald 1990). Pacific poison oak (Toxicodendron diversiloba), regenerates via seed dispersed by birds and sprouting from the root crown following fire. Birchleaf mountain mahogany is top killed by fire and sprouts via rhizomes or the root crown.
Harvesting: In studies examining oaks following harvest (McCleary, 2008) one third of the trees sprouted, however, subsequent sprout survival was significantly affected by browsing. In general, smaller diameter oaks (less than 12 inches in diameter) sprout more readily than larger oaks (Standiford, 2011) and sprouting is more common in moist environments than in dry sites (McCleary et al., 2002; McCreary 2004).
Grazing: Currently species composition and productivity of the annual-dominated grassland and understory grasses and forbs vary greatly within and between years and is greatly influenced by the timing and amount of precipitation and the amount of residual dry matter (George et al., 2001a). Grazing (or the lack of grazing) greatly influences residual dry matter (RDM) left on site. Sufficient litter or residue is required for good germination of grass species (Young et al, 1981). Besides weather and temperature, oak canopy may affect forage yield; in the drier areas of California with low oak cover, oak canopy enhances forage, and in higher precipitation areas with greater oak cover, production is reduced (Connor, 1996). Tree shading may help reduce thermal stress in cattle; as temperature increases so do water requirements (Hahn, 1985; George et al., 2015).
Diseases: There are many disease and insect pests found on California oaks. Some diseases of blue oak damage the heartwood of the trunk and large limbs (McDonald, 1990; Hickman et al., 2011). A white pocket rot (Inonotus dryophilus) causes rot in the heartwood of living oaks. The sulphur conk (Laetiporus sulphureus) causes a brown cubical rot also of the heartwood of living oaks. The hedgehog fungus (Hydnum erinaceum) and the artist’s fungus (Ganoderma applanatum) are also capable of destroying the heartwood of living oaks. A disease of blue oak roots, the shoestring fungus rot (Armillaria mellea) gradually weakens trees at the base until they fall. A white root rot (Inonotus dryadeus) also has been reported on blue oak. The fruit tree leaf roller (Archips argyrospila) may cause significant defoliation and when combined with a multi-year drought, an increase in oak tree mortality rates (USDA, 2006). Prominent diseases of foothill pine include western gall rust (Periderium harknessii) and dwarf-mistletoe (Arceuthobium occidentale) and (A. campylopodum forma campylopodum) (Howard, 1992).
Climate: In California‘s Mediterranean climate evaporative demand and rainfall are out of sync with one another (Miller et al., 2012). During peak demand in the spring, water is quickly depleted from the soil profile and grasses senesce. After that period the only moisture available to woody plants is through root access to groundwater. Groundwater has been shown to be a critical link to blue oak survival over the prolonged summer drought period (Miller et al., 2010). Extended periods of drought could slow recovery by limiting photosynthesis and affect carbon intake, hindering reproductive processes, leading to a reduction in oak seedling establishment.
The influence of climate change on vegetation has been widely debated. Some climate models indicate that decreasing precipitation and increasing temperature could result in a potential shift in the blue oak type to the north and shrinking of the overall range of the species. This change in range is thought to be a potential result of increasing moisture stress with changing climate (Kueppers, et al, 2005). Although there are many other factors that influence plant communities, climate related effects include the potential for a changed fire regime and more favorable conditions for species invasions (Stromberg, et al, 2007).
State and transition model
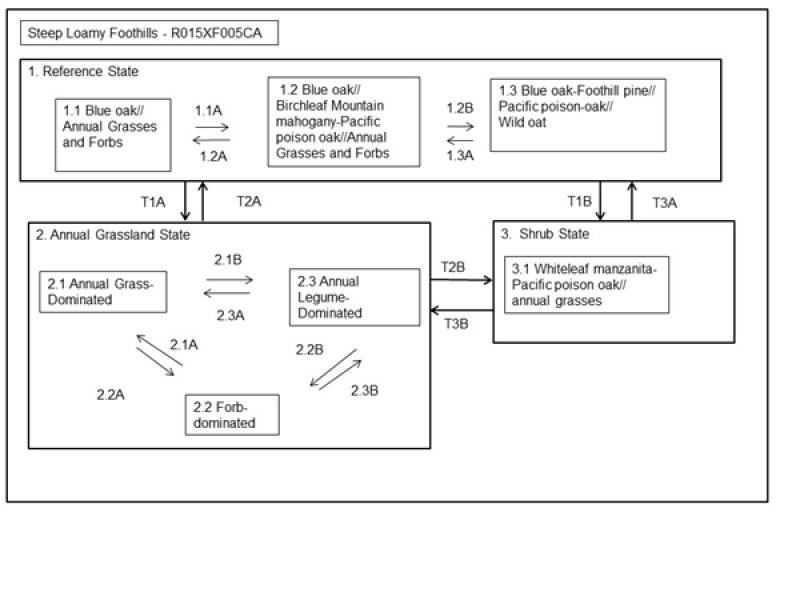
Figure 8. R015XF005CA - Steep Loamy Foothills
More interactive model formats are also available.
View Interactive Models
More interactive model formats are also available.
View Interactive Models
Click on state and transition labels to scroll to the respective text
Ecosystem states
State 2 submodel, plant communities
State 3 submodel, plant communities
State 1
Reference State
The Reference State displays the natural range of variability and ecological potential for this ecological site. States and Community Phases included in this document include those previously recognized by Fire and Resource Assessment Program (FRAP, State of California, 1998) and other entities as a result of the use of ordination software and professional consensus (Allen-Diaz et al., 1989; Vayssieres and Plant, 1998; and George et al., 1993). The reference state has three community phases: 1) Blue oak//Annual Grasses Phase: the lower elevations of this site tend to be grass and blue oak dominated. 2) Blue oak//Birchleaf mountain mahogany-Pacific poison oak//Annual Grasses and Forbs Phase: with increasing elevation and slope, tree and shrub density increases. 3) Blue oak-Foothill pine//Pacific poison oak//Wild oat: north to northeast-facing slopes with favorable moisture conditions support increased biomass. The Blue oak - Foothill pine habitat type provides important breeding habitat for a large variety of wildlife and acorns are an important food source for a variety of birds and mammals. Animal communities associated with this reference state are provided with a variety of vegetation stages and habitats. Following secondary succession annual grassland gives way to shrubs in 2-5 years; mature shrubs develop in 10 to 15 years and mature foothill pine 30 to 40 years (Mayer and Laudenslayer, 1988). Most stands of blue oak range from 80 to 100 years of age (Kertis et al., 1993), however, remnant older blue oak specimens may range to over 450 years of age (Stahle et al, 2013) in more remote or steep locations. The lack of oak regeneration and the increase in foothill pine in the understory, are of concern in the long-term existence of this habitat (Mayer and Laudenslayer, 1988). This state is relatively stable unless tree removal occurs. Blue oak contributes to soil productivity through increased soil nutrition and health under trees, and removal of trees causes changes to soil cover, water and nutrient status. Research indicates that oak removal results in a rapid decline in soil quality, including a loss in soil organic matter and nitrogen (Dalgren et al., 2003). Oaks help retain more water on site and enhance soil quality through nutrient cycling, organic matter deposition and reduced bulk density (O’Geen et al., 2010). Some deeply rooted trees and shrubs may also induce hydraulic lift, transporting water to the upper soil layers (Richards and Cadewell, 1987; Caldwell et al., 1998; Ishikawa and Bledsoe, 2000; Liste and White, 2008), supporting the development of neighboring plants. Nutrients are also concentrated around shrub bases from litter fall and from sediment capture via movement of soil particles.
Community 1.1
Blue oak//Annual Grasses and Forbs

Figure 9. Community Phase 1.1 on Contra costa soil. J.Welles, 2013
This community phase is dominated by non-native annual grasses primarily wild oat (Avena fatua) or slender oat (Avena barbata), with soft brome (Bromus hordeaceus), red brome (Bromus rubens) and a lesser amount of other forbs and herbs including common yarrow (Achillea spp.), stork’s bill (Erodium spp.), bluedicks (Dichelostemma capitatum), and knotted hedgeparsley (Torilis nodosa). The overstory is dominated by blue oak. This community phase is typically found at the lower elevational range of this ecological site.
Community 1.2
Blue oak//Birchleaf mountain mahogany-Poison oak//Annual Grasses and Forbs

Figure 10. Community Phase 1.2. J. Welles, 2014
This community phase predominantly consists of a low to moderate cover of blue oak (Quercus douglasii), a low cover of Pacific poison oak (Toxicodendron diversiloba), and low to moderate cover of birchleaf mountain mahogany, particularly on the mid slopes. Non-native annual grasses and forbs are dominated by wild oat (Avena fatua) or slender oat (Avena barbata), with a lesser amount of soft brome (Bromus hordeaceus), red brome (Bromus rubens) and the forbs longbeak or red stem stork’s bill (Erodium botrys or erodium cicutarium). Seedling establishment of birchleaf mountain mahogany is sporadic and occurs after prolonged fire-free periods (Sawyer et al., 2009). The increase in shrub cover may act to slow the water infiltration rate; improved water storage under shrubs and trees makes for plant available water later in the growing season due to decreased evaporation and shading, maintaining water longer (Gill and Burke, 1999). The increase in shrub cover causes a reduction in the amount of grass and forb cover as compared to community phase 1.1.
Community 1.3
Blue oak-Foothill pine//Poison oak //Wild oat

Figure 11. Community Phase 1.3 on Contra costa Soil. Welles, 2013

Figure 12. Community Phase 1.3 landscape on Contra costa soil. J.Welles, 2013
Community phase 1.3 represents the ecological potential and is considered the reference community phase for this site. This phase (1.3) consists of an understory of non-native annual forbs and grasses dominated by wild oat (Avena fatua) or slender oat (Avena barbata). Other grasses that may be present include soft brome (Bromus hordeaceus) and desert fescue (Vulpia microstachys). A low to moderate overstory of blue oak (Quercus douglasii) averages 15-20 percent canopy and a low cover of California foothill pine (Pinus sabiniana), averages 5 to 10 percent canopy. Shrub cover of Pacific poison oak (Toxicodendron diversiloba) averages between 5 to 20 percent cover. Whiteleaf manzanita (Arctostaphylos manzanita) is a common shrub on the higher elevations of this site. Other shrubs that may be found include birchleaf mountain mahogany, and rarely scrub oak (Quercus berberidifolia) and toyon (Heteromeles arbutifolia). Small stands of perennial native grasses that may be found in some locations on this ecological site include Chinook brome (Bromus laevipes), and California melicgrass (Melica californica). California foothill pine is regenerating in the understory and is represented in the seedling and sapling stages. In the absence of fire, foothill pine composition and cover will continue to increase at the expense of blue oak. The presence of shrubs and young pine seedlings provide ladder fuels, increasing the potential for severe fire effects to vegetation should a fire occur. Reference community phase production: Expected production is highly variable based on unfavorable normal or favorable year. Total production in a 50 to 80 percent of normal year ranges from a low of 400 to a high of 1,400 pounds per acre. The drought over the last several years has hampered efforts to sample the full range of site conditions. Historic Range Site data indicates a range of 500 to 850 lbs/acre for grass and forb production.
Figure 13. Annual production by plant type (representative values) or group (midpoint values)
Table 5. Annual production by plant type
| Plant type | Low (lb/acre) |
Representative value (lb/acre) |
High (lb/acre) |
|---|---|---|---|
| Grass/Grasslike | 318 | 530 | 848 |
| Tree | 96 | 240 | 335 |
| Forb | 12 | 120 | 168 |
| Shrub/Vine | 18 | 46 | 55 |
| Total | 444 | 936 | 1406 |
Table 6. Ground cover
| Tree foliar cover | 1% |
|---|---|
| Shrub/vine/liana foliar cover | 1% |
| Grass/grasslike foliar cover | 1% |
| Forb foliar cover | 1-2% |
| Non-vascular plants | 0-5% |
| Biological crusts | 0% |
| Litter | 61-85% |
| Surface fragments >0.25" and <=3" | 0% |
| Surface fragments >3" | 0% |
| Bedrock | 0% |
| Water | 0% |
| Bare ground | 10-27% |
Figure 14. Plant community growth curve (percent production by month). CA1501, Annual rangeland (Normal Production Year). Growth curve for a normal (average) production year resulting from the production year starting in November and extending into early May. Growth curve is for oak-woodlands and associated annual grasslands..
| Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec |
|---|---|---|---|---|---|---|---|---|---|---|---|
| J | F | M | A | M | J | J | A | S | O | N | D |
| 0 | 10 | 25 | 40 | 5 | 0 | 0 | 0 | 0 | 0 | 10 | 10 |
Figure 15. Plant community growth curve (percent production by month). CA1502, Annual rangeland (Favorable Production Year). Growth curve for a favorable production year resulting from the production year starting in October and extending through May. Growth curve is for oak-woodlands and associated annual grasslands..
| Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec |
|---|---|---|---|---|---|---|---|---|---|---|---|
| J | F | M | A | M | J | J | A | S | O | N | D |
| 0 | 10 | 20 | 30 | 25 | 0 | 0 | 0 | 0 | 5 | 5 | 5 |
Figure 16. Plant community growth curve (percent production by month). CA1503, Annual rangeland (Unfavorable Production Year). Growth curve for an unfavorable production year resulting from the production year starting in October and extending through May. Growth curve is for oak-woodlands and associated annual grasslands..
| Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec |
|---|---|---|---|---|---|---|---|---|---|---|---|
| J | F | M | A | M | J | J | A | S | O | N | D |
| 0 | 15 | 70 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 5 |
Pathway 1.1A
Community 1.1 to 1.2
Low grazing pressure and long fire-free intervals allow for shrub development on the mid-elevations of this site. Birchleaf mountain mahogany seed is spread by wind and animals, though a great deal of the seed is not viable. Pacific poison oak is spread by birds that eat the fruit and excrete the seed, and will also sprout vegetatively from the rhizomes or root crown after fire or cutting (Howard, 1994).
Pathway 1.2A
Community 1.2 to 1.1
Grazing and or browsing or repeated fire in the early stages of brush growth could slow further brush development and reduce fuel loads by reducing the seed source needed for brush re-establishment. Birchleaf mountain mahogany requires a long fire-free period to establish post-fire (Gucker, 2006) as seed viability is low and it needs moisture to germinate. Targeted grazing utilizing short periods with high stock densities in combination with other methods has met with some success in controlling brush. Goats are capable of eating spiny or poisonous plants (Popay and Field, 1996). Grazing with goats and sheep has been utilized to slow brush regrowth following mature plant removal (Stromberg, 2007). Shrub redevelopment may take 10 to 20 years. Repeated frequent fire intervals and an eventual reduction in fire intensity may deplete seed banks, kill brush and favor grasses (Stromberg, 2007). In one study, removal of shrubs by plowing with and without grazing was shown to lower the rate of infiltration (Daryanto and Eldrige, 2010) resulting in higher runoff.
Conservation practices
| Brush Management |
|---|
Pathway 1.2B
Community 1.2 to 1.3
The prolonged absence of fire (20-30 years) triggers a shift in species composition as foothill pine seedlings become established. Protection from fire and grazing results in a gradual increase in foothill pine and shrubs and contributes to increased ladder fuels and higher fuel loads.
Pathway 1.3A
Community 1.3 to 1.2
Tree removal or clearing of foothill pine results in a blue oak dominated community phase. Young regeneration of foothill pine is also susceptible to fire by virtue of its thin bark (Howard, 1992).
Conservation practices
| Forest Stand Improvement |
|---|
State 2
Annual Grassland State
Non-native grasses now have become naturalized in much of California. Introduced annual forbs and grasses have unique adaptations that give them a competitive advantage over native species. Some of these plant adaptations include high seed production, fast early season growth and the ability to set seed in drought years (Stromberg et al., 2007). Soil disturbance from burrowing animals and feral pigs continue to create new opportunities for exotic species invasion. Nutrient turnover is rapid in grassland systems and is lost via leaching, gaseous exchange and soil erosion (Stromberg et al, 2007). Because most of the nitrate that accumulates during the summer and fall is moved to seeds at senescence and the remainder is removed via rains prior to initiation of growth, little is available for later absorption by growing plants. There is a higher nutrient loss from annual systems as opposed to shrub-dominated systems (Michaelides et al., 2012) and a higher percentage of “fines” transported offsite despite similar erosion rates, according to one study. Although nutrient leaching from grassland systems is variable, nutrients that are moved beyond the shallow root systems of the annual grasses are lost to leaching. Annuals use available water primarily in the top 1 foot of soil (George et al., 2001); their shallow root structures dry out quickly during rapid spring growth and evapotranspiration quickly depletes soil moisture. Water infiltration may be more rapid in grasslands than in shrub-dominated landscapes.
Community 2.1
Annual Grass-Dominated
Common annual grasses include wild oat (Avena fatua) and slender oat (Avena barbata), soft brome (Bromus hordeaceus) and fescue (Vulpia spp.). Sufficient litter or residue is required for good germination of grass species (Young et al., 1981) and leaving greater amounts may favor grass dominance (George et al., 1985). Mulch also improves soil fertility and increases infiltration as well (Heady, 1956). Minimum residual dry matter (RDM) guidelines for dry annual grassland suggest retention of 300 to 600 pounds per acre, with greater retention as slope increases (Bartolome et al., 2002) to provide for soil and nutrient retention. Litter improves soil fertility and increases infiltration as well by providing cover during the hot summers, reducing evapotranspiration rates, leaving more moisture in the soil profile (Heady, 1956). Grasses have positive effects on soils by enhancing water percolation, aeration and carbon storage (Eviner and Chapin, 2001). Certain grasses that produce high amounts of litter such as wild oat and soft brome may attract voles and mice.
Community 2.2
Forb-Dominated
Common forb species found are longbeak stork’s bill (Erodium botrys) and redstem stork’s bill (Erodium cicutarium). Filaree years occur in low rainfall years or when residual dry matter (Bartolome et al., 2002, George et al., 1985) is low. Drought, heavy grazing and fire may all result in filaree-dominated understory. Often when a dry period follows the first rains, drought tolerant self-burial seed species are favored (Young et al, 1981). Filaree presence may be reduced if rains come early and are followed by severe drought stress (Bartolome, 1979).
Community 2.3
Legume-Dominated
Legume or clover years may be favored with early rains and regularly distributed rainfall throughout the growing season from November through April. (George et al., 1985). Dry autumn weather followed by precipitation in late fall or early winter may contribute to legume domination or clover plant community over grasses (Pitt and Heady, 1978). Rose clover (Trifolium hirtum) or minature lupine (Lupinus bicolor) are commonly found in this community phase. Legumes enhance nitrogen availability, but may increase leaching and gopher activity (Eviner and Chapin, 2001).
Pathway 2.1A
Community 2.1 to 2.2
Filaree years are triggered in low rainfall years or when residual dry matter (Bartolome et al., 2002, George et al., 1985) is low. Often when a dry period follows the first rains, drought-tolerant self-burial seed species, like filaree, are favored (Young et al., 1981) and the deep taproot of filaree supplies water to the plant (Pitt and Heady 1978). Filaree presence may be reduced if rains come early and are followed by severe drought stress (Bartolome, 1979). Filaree provides forage early in the growing season but rapidly disintegrates after maturity and often leaves a lot of exposed soil (Pitt and Heady, 1978). Successive droughts could lead to erosion especially after a filaree year.
Pathway 2.1B
Community 2.1 to 2.3
Legume or clover years may be favored with early rains and regularly distributed rainfall throughout the growing season from November through April. (George et al., 1985).
Pathway 2.2A
Community 2.2 to 2.1
Annual grass years occur when precipitation is high or with late spring rains (George et al, 1985). Annual grasses are shallow-rooted species that require a continual supply of moisture for growth (Barbour and Major, 1977).
Pathway 2.2B
Community 2.2 to 2.3
Legume domination or clover years may be favored with early rains and adequately spaced rainfall thereafter (George et al, 1985). Low mulch cover may allow for increased development of legumes (Heady, 1956).
Pathway 2.3A
Community 2.3 to 2.1
Annual grass years occur when precipitation is high or with late spring rains (George et al, 1985). Annual grasses are shallow-rooted species that require a continual supply of moisture for growth (Barbour and Major, 1977). Sufficient litter or residue is required for good germination of grass species (Young et al., 1981) and leaving greater amounts may favor grass dominance (George et al., 1985). Litter also improves soil fertility and increases infiltration as well by providing cover during the hot summers, reducing evapotranspiration rates, leaving more moisture in the soil profile (Heady, 1956).
Pathway 2.3B
Community 2.3 to 2.2
Filaree years are triggered in low rainfall years or when residual dry matter (Bartolome et al., 2002; George et al., 1985) is low. Often when a dry period follows the first rains, drought-tolerant self-burial seed species, like filaree, are favored (Young et al., 1981) and the deep taproot of filaree supplies water to the plant (Pitt and Heady 1978). Filaree presence may be reduced if rains come early and are followed by severe drought stress (Bartolome, 1979).
State 3
Shrub State
Upper drier backslope positions tend to be droughty and support more whiteleaf manzanita in the shrub layer. As development of the shrub community progresses after fire, inter-shrub native and non-native herbaceous vegetation decreases, and less understory vegetation is remaining. In a shrub state available water may be present later in the growing season due to decreased evaporation and shading, maintaining moisture longer than under just grasses alone (Gill and Burke, 1999). Deeply rooted shrubs may also induce hydraulic lift, transporting water to the upper soil layers (Richards and Cadewell, 1987). Nutrients are also concentrated around shrub bases from litter fall and from sediment capture via movement of soil particles.
Community 3.1
Pacific Poison Oak-Whiteleaf Manzanita//Annual Grasses
Post-fire recovery is from residual species that sprout or from dormant seed banks. Whiteleaf manzanita produces abundant seed that is stored in a dormant condition until stimulated to germinate by fire (Abrahamson, 2014). Initially, there may be a large amount of manzanita seedlings that are later greatly reduced due to competition or drought (Abrahamson, 2014). This shrub is sporadically able to regenerate without fire, though produces isolated individuals and grows best in open conditions. Pacific poison oak sprouts from rhizomes (Howard, 1994) or the root crown and may also reproduce by layering. Poison oak seeds are also spread by birds. While not well documented, repeated fire could facilitate the presence and possible expansion of Pacific poison oak through re-sprouting of existing plants or through seed transported by birds and other mammals (Howard, 1994). The amount of Pacific poison oak present varies depending on the intensity, frequency and timing of fire. Increases in Pacific poison oak often occur after disturbance in the short-term and generally decreases with increasing shade and dominance of higher plants. Other shrubs that may be found on lower elevations of this ecological site that could be a minor part of this community include birchleaf mountain mahogany, scrub oak (Quercus berberidfolia) and toyon (Heteromeles arbutifolia). They are top killed and resprout following cutting or burning from rhizomes or the rootcrown.
Transition 1B
State 1 to 2
As community phases change over time within the Reference State, fire regimes may shift from low intensity fires towards more mixed severity and replacement fires due to the presence of ladder fuels and several missed fire cycles. A moderate to severe fire that kills blue oak and foothill pine is a trigger that could result in reaching a Threshold (T1B) causing a transition to a shrub/annual grass and forb state (State 3). Some smaller blue oak may survive, but larger oaks would likely have low sprouting and reduced or eliminated re-seeding capacity. Fuel load, season of burn and fire frequency interact to affect the amount of damage and mortality response (Swiecke et al., 1997). Pacific poison oak re-sprouting is stimulated by fire (Howard, 1994). Shrub species such as whiteleaf manzanita that have seed stored in the soil has abundant germination following fire (Abrahamson, 2014). Other shrubs that may be present in lesser amounts such as scrub oak (Quercus beberidifolia), birchleaf mountain mahogany (Cercocarpus montanus var glaber) and toyon (Heteromeles arbutifolia) sprout from the root crown following cutting or fire. Competition from shrubs for moisture and growing space may also limit re-establishment of blue oak.
Transition 1A
State 1 to 2
Triggers that limit blue oak reproduction and/or survival (intensive prolonged grazing) in combination with fire or mechanical removal and prolonged drought that cause mortality or unfavorable conditions for sprouting result in a Threshold (T1A), transitioning to an annual grassland state ( State 2). Natural regeneration seldom occurs further away than 100 feet from an existing oak tree canopy, and stump sprouting is likely to be very limited in mature trees in a low rainfall area (FRAP, State of California, 1998). Blue oak seedlings may be less adapted to drought conditions, affecting their growth and survival (Grünzweig et al., 2008). One study (McCreary et al., 2006) found sprout survival 19 years following harvest was 28 percent and noted that sprouting would be expected to decline with any subsequent harvest. Removal of blue oaks has been found to reduce soil productivity due to decreased soil nutrition from tree litter (Dalgren et al., 2003), and increase the potential for erosion due to a decline in soil porosity and increased bulk density from organic matter losses (Dalgren et al., 1997). Erosion losses also may reduce productivity by changing the water-holding capacity of the soil and the thickness of the root zone (Elliot et al., 1998).
Transition T2A
State 2 to 1
Removal of oak trees and subsequent soil erosion may change runoff and infiltration rates making restoration efforts difficult without significant inputs. Planting success might be achieved, especially on the low elevation sites, through planting acorns or seedlings and tending young trees using tested methods (McCreary, 2004) and by protecting young oaks from grazing by utilizing fencing. Blue oak saplings take approximately 10 to 30 years to reach a height of about 4.5 feet (Swiecki, 1998).
Transition T2B
State 2 to 3
Shrub recruitment has increased in the absence of periodic fire or grazing in some foothill environments (Duncan, 1987). Though unlikely to take over large areas with annual grasses, drier mid to high elevation sites would be most susceptible to shrub encroachment.
Transition T3A
State 3 to 1
Over a period of time, blue oak and California foothill pine may initiate limited re-establishment through sprouting and from adjacent seed sources. Pioneer reproduction of oak in rangelands is uncommon (Swiecki and Bernhardt, 1998). While studies are lacking on growth of the species found on this site, mature shrubs can take 10-20 years to develop, mature pine 30 to 50 years, and blue oak 40 to 100+ years (Mayer and Laudenslayer, 1998) to reach maturity.
Transition T3B
State 3 to 2
Short fire return intervals act to kill shrubs and reduce the seed bank, and promote herbaceous annuals, creating a grass/herbaceous mixture. Repeated grazing would also contribute to a reduction in shrub cover. Continued high fire frequency can convert shrubs to an annual grassland dominated by non-native and native grasses and forbs (Keeley, 2002).
Additional community tables
Table 7. Community 1.3 plant community composition
| Group | Common name | Symbol | Scientific name | Annual production (lb/acre) | Foliar cover (%) | |
|---|---|---|---|---|---|---|
|
Forb
|
||||||
| 1 | Annual Herbs | 58–72 | ||||
| springbeauty | CLAYT | Claytonia | 10–23 | 1–5 | ||
| false carrot | YAMI | Yabea microcarpa | 13–21 | 1–3 | ||
| knotted hedgeparsley | TONO | Torilis nodosa | 10–17 | 1–2 | ||
| rose clover | TRHI4 | Trifolium hirtum | 0–11 | 0–1 | ||
| pincushionplant | NAVAR | Navarretia | 5–8 | 1–2 | ||
| 5 | Perennial Herbs | 44–83 | ||||
| straightbeak buttercup | RAOR3 | Ranunculus orthorhynchus | 26–47 | 2–8 | ||
| Ithuriel's spear | TRLA16 | Triteleia laxa | 2–15 | 1–3 | ||
| onion | ALLIU | Allium | 8–11 | 1–2 | ||
| bedstraw | GALIU | Galium | 3–6 | 1–2 | ||
| sanicle | SANIC | Sanicula | 2–4 | 1–2 | ||
|
Grass/Grasslike
|
||||||
| 2 | Grasses | 430–616 | ||||
| wild oat | AVFA | Avena fatua | 424–616 | 15–25 | ||
| Chinook brome | BRLA3 | Bromus laevipes | 0–106 | 0–6 | ||
| soft brome | BRHO2 | Bromus hordeaceus | 0–60 | 1–15 | ||
| small fescue | VUMI | Vulpia microstachys | 0–60 | 0–6 | ||
| California melicgrass | MECA2 | Melica californica | 0–8 | 0–1 | ||
|
Shrub/Vine
|
||||||
| 3 | Shrubs | 46–96 | ||||
| birchleaf mountain mahogany | CEMOG | Cercocarpus montanus var. glaber | 10–53 | 3–8 | ||
| Pacific poison oak | TODI | Toxicodendron diversilobum | 10–20 | 5–20 | ||
| scrub oak | QUBE5 | Quercus berberidifolia | 0–20 | 0–14 | ||
| whiteleaf manzanita | ARMA | Arctostaphylos manzanita | 1–3 | 1–5 | ||
|
Tree
|
||||||
| 4 | Trees | 78–330 | ||||
| blue oak | QUDO | Quercus douglasii | 61–229 | 10–25 | ||
| California foothill pine | PISA2 | Pinus sabiniana | 9–105 | 5–10 | ||
Interpretations
Animal community
Oak savannas and woodlands may provide essential habitat elements for a variety of wildlife species. The blue oak-gray pine community is preferred habitat for black-tailed deer, California quail, and mourning dove and the seeds are eaten by various birds and rodents including Scrub jay, acorn woodpecker, and squirrels are major seed consumers (Howard, 1992).
Due to the natural mosaic of grassland, shrubs and trees, a variety of micro-habitats are provided for wildlife. The increased shrub cover and structure provided in Plant Communities 1.2 and 1.3 provide additional habitat elements necessary for a variety of birds and animals. In one study in central California, habitat elements that included shrubs, grass and down wood were positively associated with the abundance of small mammals, and shrub cover and litter weight with abundance of birds and reptiles (Tieje, Vreeland, Siepel and Dockter, 1997). Shrubs provide thermal hiding and escape cover for many birds and mammals, including deer and rabbits (Fryer, 2006).
Of the 632 terrestrial vertebrates (amphibians, reptiles, birds, and mammals) native to California, over 300 species use oak woodlands for food, cover and reproduction, including at least 120 species of mammals, 147 species of birds and approximately 60 species of amphibians and reptiles (Tietje et al. 2005). Common species on this site include Beechey ground squirrels (Spermophilus beecheyi), Botta pocket gopher (Thomomys bottae mewa), and Audubon cottontail (Sylvilagus audubonii vallicola). The rich rodent and lagomorph population is an important food source for common predators including: bobcat (Lynx rufus californicus), coyote (Canis latrans) and the Pacific rattlesnake (Crotalus viridis oreganus).
Other wildlife species found in oak woodland include several important game animals, such as mule deer (Odocoileus hemionus), California quail (Callipepla californica), and the "re-introduced" wild turkey (Meleagris gallopavo) contribute to California’s economy through revenues from recreational hunting (Garrison and Standiford, 1997).
Birds can serve as “focal species” in that their requirements define spatial attributes, habitat characteristics and management regimes for a healthy system (Zack, 2002). Essential habitat elements include large oak trees with associated cavities and acorns, snags, shrubs, grasses and forbs, brush piles and water. Oak woodlands are important over-wintering environments for large numbers of neotropical migratory birds such as flycatchers, vireos, and warblers. Acorn woodpeckers (Melanerpes formicivoris) and western scrub jays (Aphelocoma californica) forage heavily on acorns, and oak titmice (Baeolophus inornatus), western bluebirds (Sialia Mexicana) and tree swallows (Tachycineta bicolor) nest in the cavities of oaks.
The numerous diseases that affect the heartwood of oak boles and limbs create important cavities for a variety of tree dwellers. Mistletoe in oaks (Phorodendron villosum) has berries that are attractive to birds that eat and excrete the live seeds which then stick and create new plants (Perry and Elmore, 2006).
Different age classes of blue oak (seedlings, sapling and young trees) are represented on these north slope ecological sites. Shade ameliorates soil temperatures for seedling establishment and the animals that utilize this site. Birds perch in the trees and contribute to shrub dispersal (Block, 1990). Forage and cover available for birds and other animals on these sites is low to moderate.
Grazing and Browsing
The primary limitation for livestock production on this site is lack of water sources during most of the year.
The quality of range forage varies with plant species, season, location, and range improvement practices. Range forage is optimal for livestock growth and production for only a short period of the year (George, 1993).
The browse value of common oak woodland species is listed in Sampson and Jespersen (1963). Manzanita berries are utilized by a number of birds and small animals (Stuart and Sawyer, 2001). Young oak seedlings leaves are browsed by deer and cattle.
Acorns are eaten by at least a dozen species of songbirds, several upland game birds, rodents, black-tailed deer, feral and domestic pig, and all other classes of livestock (Adams et al., 1992; Duncan and Clawson 1980; Sampson and Jespersen 1963). Acorns are a critical food source for deer, which migrate from high-elevation dry summer ranges to blue oak woodland for fall and winter forage (Burns and Honkala 1990). Deer, rodents and rabbits browses blue oak and contributes to poor survival of oak natural regeneration.
Hydrological functions
The watersheds associated with these sites are drained by intermittent streams that only flow during the wet season. In dry years these intermittent streams may not flow at all. Runoff on these soils is rapid and soil erosion hazard is high.
Recreational uses
Bird watching, hunting, camping, horseback riding, all-terrain vehicle riding, and hiking in spring are common recreational pursuits.
Wood products
Firewood cutting of blue oak, once prevalent, has decreased with increased public awareness of poor blue oak regeneration.
Other products
Native Americans have historically used and managed the blue oak woodlands for food and fiber. The gathering of native plants such as bulbs and corms, grasses and brush for food, medicine and crafts is still practiced today (Anderson, 2006). These gathering methods sustained local plant populations and promoted plant diversity.
Other information
Natural regeneration of blue oaks may be limited on some sites and because of a number of factors that limit seed germination, seedling establishment and survival to the tree stage. Competition for soil moisture from the understory annual plants, acorn and seedling damage by rodents, livestock grazing and changed fire regimes are important factors that can reduce blue oak regeneration. McCreary (2001) provides an extensive review of oak regeneration problems and practices on California’s oak woodlands. Young oak seedlings are especially susceptible to mortality during the early years. One study indicates blue oak seedlings that reach 10 years of age are more likely to survive compared to newly germinated seedlings (Philips et al, 2007). Protection of saplings from grazing pressure utilizing tree shelters (McCreary and George, 2005) or exclosures (Philips, et al 2007) has been shown to be an effective management technique to aid survival and accelerate growth of young seedlings. Supplemental irrigation may be necessary to ensure survival on some sites.
Native Grass Restoration: Native perennial grasses may occur on this ecological site in very small amounts. There is no known practice or group of practices that can successfully restore native grasses on this ecological site.
Soils: The capacity of the soil to function is affected by dynamic soil properties (Tugel and Brown, 2001). “Dynamic properties include organic matter, soil structure, infiltration rate, bulk density, and water and nutrient holding capacity” (Soil Quality.Org, 2012). Soil resistance is defined as: "the capacity of a soil to continue to function without change throughout a disturbance" and soil resilience is defined as "the capacity of a soil to recover its functional and structural integrity after a disturbance" (Seybold et al. 1999). “Changes in those properties depend both on land management practices and the inherent properties of the soil” (Soil Quality.Org, 2012).
These soils have some properties that have a high resistance to disturbance; for example they have a moderate volume (moderate depth) to absorb and buffer compaction (average soil depth is about 35 inches). Infiltration and water and nutrient holding capacity are enhanced by the strong structure found in the well developed soil profile.
Supporting information
Inventory data references
Ecological Site Inventory Plots:
- 9 traverses with soil pits
- 4 Line intercept transects with soil pits
- 7 double sampling plots
Range 417s - #14, 1981
Type locality
| Location 1: Colusa County, CA | |
|---|---|
| Township/Range/Section | TT15N RR5W S11 |
| UTM zone | N |
| UTM northing | 551859.6 |
| UTM easting | 4335528.6 |
| General legal description | Location approximately 1.3 miles east of Bear Valley Road. |
Other references
Abrahamson, I. 2014. Arctostaphylos manzanita. In: Fire Effects Information System, [Online]. U.S. Dep. of Agric., For. Serv., Rocky Mountain Res. Sta., Fire Sci. Lab. (Producer). Available: http://www.fs.fed.us/database/feis/ [2014, July 21].
Adams, Theodore E., P. B. Sands, W.H. Weitkamp, and N. K. McDougald. 1992. Oak seedling establishment on California rangelands. J. Range Manage. 45: 93-98.
Allen-Diaz, Barbara, R. R. Evett, B. A. Holzman, and A. J. Martin. 1989. Report on Rangeland Cover Type Descriptions for California Hardwood Rangelands. Forest and Rangeland Resources Assessment Program, Calif. Dep. of Forestry and Fire Protection, Sacramento, Calif. 318 pgs.
Anderson, M.K. 2005. Tending the Wild, Native American Knowledge and the Management of California’s Natural Resources. University of California Press.
Arno, S. H. and S. Allison-Bunnell. 2002. Flames in Our Forest. Island Press. 227 pgs.
Baldocchi, D.D., and L. Xu. 2007. What limits evaporation from Mediterranean oak woodlands – The supply of moisture in the soil, physiological control or the demand by the atmosphere? Elsevier Ltd. Doi:10.1016/j.advwatres.2006.06.013. Advances in Water Resources. 30 (2007) 2113-2122. www.elsevier.com/locate/advwatres
Baldocchi, D.D., Xu, L., and N. Kiang. 2004. How plant functional type, seasonal drought and soil physical properties alter water and energy fluxes on oak-grass savanna and an annual grassland. Agr. and For. Meterology. Vol.123, issues 1-2, pg.13-39.
Barbour, M.G., and J. Major. Terrestrial Vegetation of California. 1977. John Wiley and Sons, Inc.
Bartolome, J.W. 1979. Germination and Seedling Establishment in California Annual Grassland. Journal of Ecology. Vol.67, No. 1, 7 pgs.
Bartolome, J. W. 1987. California grassland and oak savannah. Rangelands 9:122-125.
Bartolome, J.W., W.F. Frost, N.K. McDougald and M. Connor. 2002. California guidelines for residual dry matter (RDM) management on coastal and foothill annual rangelands. Rangeland Monitoring Series. Publ. 8092, Div. of Agr. and Nat Res., Univ. of Calif. 8pp.
Blackburn, T.C. and K. Anderson. 1993. Before The Wilderness: Environmental Management By Native Californians. Ballena Press, Menlo Park, CA.
Bolsinger, C. L. 1988. The hardwoods of California’s timberlands, woodlands, and savannas. Res. Bull. PNW-RB-148. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 148 p.
Burcham, L. T. 1957. California Rangeland. Div. Forestry, Sacramento, Calif. 261 pgs.
Burns, Russell M. and B. H. Honkala. 1990. Silvics of North America (Vol. 2): Hardwoods. Agric. Handbook 654. USDA Forest. Service, Washington D.C. 877 p.
Conner, M. 1991. http://ucanr.edu/sites/oak_range/Oak_Articles_On_Line/Factors Affecting Annual Forage Yield in Oak Woodlands.
Corbin, J. D. and C. M. D’Antonio. 2004. Competition between native perennial and exotic annual grasses: Implications for an historical invasion. Ecology 85:1273-1283.
Dahlgren R., Horwath W., Tate K., Camping T. 2003. Blue oak enhance soil quality in California oak woodlands. Calif. Agr. 57(2):42-47. DOI: 10.3733/ca.v057n02p42
Dahlgren, R.A., K.W. Tate, D. Lewis, D.R. Atwill, J.M. Harper, and B. Allen-Diaz. 2001. Watershed research examines rangeland management effects on water quality. CALIFORNIA AGRICULTURE, VOL. 55, No. 6.
Dahlgren, R.A., Singer, M. J., and X. Huang. 1997. Oak tree and grazing impacts on soil properties and nutrients in a California oak woodland. In: Biogeochemistry 39:45-64. Kluwer Academic Publishers.
Daniel, J.A., K. Potter, W. Altom, H. Aljoe, and R. Stevens. 2002. Long-Term Grazing Density Impacts on Soil Compaction. Transactions of the ASAE, 45:1911-1915.
Daryanto, S. and D.J. Eldridge. 2010. Shrub removal and grazing alter the spatial distribution of infiltrability in a shrub-encroached woodland. In: Proceedings of the 16th Biennial Conference of the Australian Rangeland Society, Bourke (Eds. D.J. Eldridge and C. Waters) (Australian Rangeland Society:Perth)
D’Antonio, C.D, S. Bainbridge, C. Kennedy, J. Bartolome, S. Reynold. 2006. Ecology and Restoration of California Grasslands with special emphasis on the influence of fire and grazing on native grassland species. University of California, Berkeley. Department of Integrative Biology.
Duncan, D. A. and W.J. Clawson. 1980. Livestock utilization of California’s oak woodlands. In: Plumb, Timothy R., (technical coordinator). Proceedings of the symposium on the ecology, management, and utilization of California oaks. Gen. Tech. Rep. PSW-44. U.S. Dep. of Agr., For. Serv. Pacific Southwest Forest and Range Exp. Sta., Berkeley, CA. Pgs. 306-313.
Eliot, W.J., Page-Dumroese, D.S., and P.R. Robichaud. 1998. The effect of forest management on erosion and soil productivity. In: Lal, R., Soil quality and erosion. Boca Raton, FL: St. Lucie Press, 195-209.
Eviner, V.T. and F.S. Chapin III, 2001. Plant species provide vital ecosystem functions for sustainable agriculture, rangeland management and restoration. California Agriculture, Volume 55, Number 6. Pgs. 54-59.
Fryer, J. L. 2007. Quercus douglasii. In: Fire Effects Information System, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer). Available: http://www.fs.fed.us/database/feis/ [2013, February 19].
Garrison, B.A., and R.B. Standiford, 1997. A Post-Hoc Assessment of the Impacts to Wildlife Habitat from Wood Cutting in Blue Oak Woodlands in the Northern Sacramento Valley. USDA Forest Service Gen. Tech. Rep. PSW-GTR-160.
George, M., Harper, J., Davy, J., and T. Bechetti. Livestock Production. http://californiarangeland.ucdavis.edu/Livestock_Production/ accessed May 1, 2015.
George, M., J. Clawson, J. Menke, and J. Bartolome. 1985. Annual grassland Forage Productivity. 1985. In: Rangelands 7(1), 3 pgs.
George, M., J. Bartolome, N. McDougald, M. Connor, C. Vaughn and G. Markegard. 2001a. Annual Range Forage Production. ANR Publ. 8018, Div. of Agric. And Nat. Res., Univ. of Calif., Oakland, Calif. 9 pgs.
George, M., G. Nader, N. McDougald, M. Connor, and B. Frost. 2001b. Annual Rangeland Forage Quality. ANR Publ. 8022, Div. of Agric. And Nat. Res., Univ. of Calif., Oakland, Calif. 13 pgs.
George, M.R., J.R. Brown and W.J. Clawson. 1992. Application of non-equilibrium ecology to management of Mediterranean grasslands. J. Range Manage. 45: 436-440.
George, M. 1993. ESDs https://esis.sc.egov.usda.gov/
Griffin, D., and K. J. Anchukaitis (2014), How unusual is the 2012–2014 California drought?, Geophys. Res. Lett., 41, 9017–9023, doi:10.1002/2014GL062433.
Gill, R. A., and I.C. Burke. 1999. Ecosystem consequences of plant life form changes at three sites in the semi-arid United States. Oecologia (1999) 121:551-563
Grünzweig, J.M., Carmel, Y., Riov, J., Sever, N., McCreary, D.D., and C.H. Flather, 2008. Growth, resource storage, and adaptation to drought in California and eastern Mediterranean oak seedlings. Can. J. For. 38: 331-342 (2008) doi:10.1139/X07-152
Hahn, G.L. 1995. Weather and Climate Impacts to Beef Cattle. USDA, Ag. Res. Sta., Lincoln, Nebraska. Published in Beef Research Program Progress Report (1985) No. 2: 85-89
Harper, J.M., Standiford, R.B., and J. Leblanc. 1991. http://ucanr.edu/sites/oak-range/Oak_Articles_On_Line/Oaks and Climatic Factors/The Effect of Drought on California Oaks.
Heady, H. F. 1956. Changes in a California Annual Plant Community Induced by Manipulation of Natural Mulch. In Ecology, Vol.7, No.4. pgs. 798-812.
Herman, D.J., Halverson, L.J., and M.K. Firestone, 2003. Nitrogen Dynamics in an Annual Grassland: Oak Canopy, Climate and Microbial Population Effects. Ecological applications, 13(3) 2003, pp. 593-604.
Hickman, G.W., E.J. Perry, and R.M. Davis. 2011. Wood Decay Fungi in Landscape Trees. University of California. Integrated Pest Management Program. Agriculture and Natural Resources. Pest Notes 74109.
Howard, Janet L. 1992. Pinus sabiniana. In: Fire Effects Information System, [Online].
U.S. Dep. of Agric., For. Ser., Rocky Mountain Res. Sta., Fire Sciences Lab. (Producer). Available: http://www.fs.fed.us/database/feis/ [2015, April 30].
Keeley, J.E. 2002. Native American impacts on fire regimes of the California coastal ranges. Jor. of Biogeography, 29, 303-320.
Kertis, J.A., R. Gross, D.L. Peterson, M.J. Arbaugh, R.B. Standiford, D.D.
McCreary.1993. Growth trends of Blue oak (Quercus douglasii in California. Can. Jor. For. Res. 23:1720-1724.
Kreitlow, K.W. and R.H. Hart.1974. Plant morphogenesis as the basis for scientific management of range resources: Agricultural Research Service, U.S. Dept. of Agriculture. Proceedings of the workshop of the United States-Australia Rangelands Panel, Berkeley, California, March 29-April 5, 1971 (Google eBook)
Kueppers, L.M., M.A. Synder, L.C. Sloan, E.S. Zavaleta, B. Fulfrost. 2005. Modeled regional climate change and endemic oak ranges. Proceedings of the National Academy of Sciences of the United States of America. www.pnas.org_cgi_doi_10.1073_pnas.0501427102. Vol. 102. No.45.
Mayer K. E., and W. F. Laudenslayer. (Eds.) 1988. A guide to wildlife habitats of California. California Dept. of Forestry and Fire Protection, Sacramento.
McCreary, D.D. and M. George. 2005. Managed Grazing and Seedling Shelters Enhance oak Regeneration on Rangelands. http://californiaagriculture.ucop.edu/
McCreary, D.D. Fire in California’s Oak Woodlands. 2005. http://danr.ucop.edu/ihrmp
McCreary, D. D. 2001. Regenerating rangeland oaks in California. ANR Publ. 21601, Div. of Agric. And Nat. Res., Univ. of Calif., Oakland, Calif. 62 pgs.
McCreary, D.D., Tietje, W.D., and W. Frost. 2002. Stump sprouting of Blue Oaks Ten Years after Harvest. USDA For. Serv., Gen.Tech.Rep. PSW-GTR-184. 8 pgs.
McCreary, D.D., Tieje, W., and W. Frost. 2008. Stump Sprouting 19 Years after Harvest. Gen. Tech. Rep. PSW-GTR-217.
McClaran, M.P. 1986. Age structure of Quercus douglasii in relation to livestock grazing and fire. Ph.D. Dissertation. Univ. of Calif., Berkeley. 119 pp.
McDougald, N.K. W.E. Frost, and W.J. Clawson. 1991. Estimating the cost of replacing forage losses on annual rangeland. Leaflet 21494. Division of Agric. and Nat. Res., Univ. of Calif., Oakland, Calif.
McDonald, P. M. 1990. Quercus douglasii Hook & Arn. Blue oak. In: Burns, Russell M.; Honkala, Barbara H., tech. coords. Silvics of North America. Vol.2. Hardwoods. Agric. Handb. 654. Washington, DC: U.S. Department of Agriculture, Forest Service: 631-639.
Mensing, S. A. 1992. The impact of European settlement on blue oak (Quercus douglasii) regeneration and recruitment in the Tehachapi Mountains, California. Madrono. 39: 36-46.
O’Geen. A.T., Dalgren, R.A., Swarowsky, A., Tate, K.W., Lewis, D.J., and M.J. Singer. 2010. Research connects soil hydrology and stream water chemistry in California oak woodlands. CA. Agric. Vol. 64. No.2. http://escholarship.org/uc/item/34x9m71j
Parsons, D.J and T.K. Stohlgren. 1989. Effects of varying fire regimes on annual grasslands in the southern Sierra Nevada of California. Madroño, 36:154-168.
Pavlik, B.M., P.C. Muick, S. Johnson, and M. Popper. 1991. Oaks of California. Cachuma Press, Inc. Los Olivos, Calif. 184 pgs.
Perry, E.J. and C.L Elmore. 2006. University of California. Mistletoe. Integrated Pest Management Program. Agriculture and Natural Resources. Pest Notes 7437.
Philips, R.L., N.K. McDougald, D. McCreary, and E.R. Atwill. 2007. Blue oak seedling age influences growth and mortality. California Agriculture, Vol.61, No.1.
Philips, R.L., N.K. McDougald, E.R. Atwill, and D. McCreary. 2007. Exclosure size affects young blue oak seedling size. California Agriculture, Vol.61, No.1. 4pgs.
Pickett, S.T.A., and P.S. White. 1985. The Ecology of Natural Disturbance and Patch Dynamics. Academic Press. 472 Pgs.
Pitt, M. D. and H. F., Heady. 1978. Responses of Annual Vegetation to Temperature and Rainfall Patterns in Northern California. Ecology, Vol.59, No.2, pgs. 336-350.
Popay, I. and R. Field. 1996. Grazing Animals as Weed Control Agents. Weed Technolgy. Vol10, No.1, pp. 217-213.
Purcell, K.L., Stephens, S.L., 2005. Changing fire regimes and the avifauna of
California oak woodlands. Studies in Avian Biology 30, 33–45.
Quinn, R.D. and S.C. Keeley. 2006. Introduction to California Chapparal. University of California Press. 322 pgs.
Richards, J. H., and M.M. Cadewell, 1987. Hydraulic lift: Substantial nocturnal water transport between soil layers by Artemesia tridentada roots. Oecologia (Berlin) (1987) 73:486-489.
Sampson, A. W. and B. S. Jespersen. 1963. California range brushlands and browse plants. Univ. of Calif. Div. of Agr. Sci., Berkeley, CA. 162 pgs.
Sawyer, J. O., Keeler-Wolf, T., and J. M. Evens. 2009. A Manual of California Vegetation, Second Edition. California Native Plant Society Press. Sacramento, CA.
Stahle D.W., R.D. Griffin, D.M. Meko, M.D. Therrel, J.R. Edmondson, M.K. Cleaveland, L.N. Stahle, D.J. Burnette, J.T. Abatzoglou , K.T. Redmond, M.D. Dettinger, and D.R Cayan. 2013. The Ancient Blue Oak Woodlands of California: Longevity and Hydroclimatic History. Earth Interactions Volume 17. Paper No. 12.
Standiford, R.B., McCreary, D.D., Barry, S.J., Forero, L.C., 2011. Blue oak stump sprouting evaluated after firewood harvest in northern Sacramento Valley. Ca. Agric. Vol. 65, No. 3. http:/scholarship.org/us/item/5pv6j50d
State of California, Cal. Dep. For. and Fire Protection (CALFIRE) FRAP Publication: State and Transition Models for California’s Rangelands. http://frap.fire.ca.gov/projects/hardwood_expert/building_state/overview2.htm
Stephens, S.L. Fire history of mixed oak-pine forest in the foothills of the Sierra Nevada, El Dorado County, Calif. In: N.H. Pillsbury, J. Verner, and W.D. Tietje (Ed). 1997. Proceedings, Symposium on Oak Woodlands: Ecology, Management, and Urban Interface Issues. USDA Forest Service GTR-PSW GTR-160.
Stromberg, M.R., J.D. Corbin, C.M. D’Antonio. 2007. California Grasslands: Ecology and Management. University of California Press, Berkeley and Los Angeles, CA. 390 pgs.
Stuart, J.D. and J.O. Sawyer, 2001. Trees and Shrubs of California. University of California Press. 467 pgs.
Swiecki, T.J., and E. Bernhardt. 1998. Understanding Blue Oak Regeneration. Fremontia 26 (1):19-26
Swiecki, T.J., Bernhardt, E.A., and C. Drake. 1997 Stand-level Status of Blue Oak Sapling Recruitment and Regeneration. Proceedings, Symposium on Oak Woodlands: Ecology, Management, and Urban Interface Issues. USDA For. Serv. Gen. Tech. Rep. PSW-GTR-160. 147-156
Sweitzer, Rick A. and Dirk H. Van Vuren. 2002. Rooting and Foraging Effects of Wild Pigs on Tree Regeneration and Oak Survival in California’s Oak Woodland Ecosystems. USDA Forest Service Gen. Tech. Rep. PSW-GTR-184.
Tietje, W.D., J.K. Vreeland, N.R. Siepal, and J.L. Dockter. 1997. Relative Abundance and Habitat Associations of Vertebrates in Oak Woodlands in Coastal Central California. USDA Forest Service Gen. Tech. Rep. PSW-GTR-160. 1997.
Tietje, W., K. Purcell and S. Drill. Oak woodlands as wildlife habitat. In: Giusti, Gregory A., Douglas D. McCreary, and Richard B. Standiford (Ed). 2005. A Planner’s Guide for Oak Woodlands, 2nd Ed. ANR Publ. 3491, Div. of Agric. and Nat. Res., Univ. of Calif., Oakland, Calif. pp 15-31.
U.S. Dep. Agric., Nat. Res. Cons. Serv., 1999. Soil taxonomy. A Basic System of Soil Classification for Making and Interpreting Soil Surveys. Agric. Handbook 436.
University of California Agriculture ~ California Agriculture ~ January-March 2007 http://calag.ucop.edu/0701JFM/resup02.html
University of California. UC Davis. California rangelands.
http://californiarangeland.ucdavis.edu/Weeds/Weeds.htm
University of California Agriculture ~ California Agriculture ~ January-March 2007. Oaks Research Update. http://calag.ucop.edu/0701JFM/resup02.html
University of California. 2010. Sudden Oak Death. UC Statewide Integrated Pest Management Program. Agriculture and Natural Resources. http://www.ipm.ucdavis.edu/PMG/PESTNOTES/pn74151.html
Vayssieres M.P. and R.E. Plant. 1998. Identification of vegetation state-and-transition domains in California's hardwood rangelands. FRAP Publication, California Department of Forestry and Fire Protection. Sacramento, CA.
Young, J.A., Evans, R.A., Raguse, C.A., and J. R. Larson, 1981. Germinable seeds and periodicity of germination in annual grasslands. Hilgardia 49:1-37.
Zack, S. 2002. Oak Woodland Bird Conservation Plan, A Strategy for Protecting and Managing Oak Woodland Habitats and Associated Birds in California. California Partners in Flight. V.2.0
Contributors
Judith Welles
Rangeland health reference sheet
Interpreting Indicators of Rangeland Health is a qualitative assessment protocol used to determine ecosystem condition based on benchmark characteristics described in the Reference Sheet. A suite of 17 (or more) indicators are typically considered in an assessment. The ecological site(s) representative of an assessment location must be known prior to applying the protocol and must be verified based on soils and climate. Current plant community cannot be used to identify the ecological site.
| Author(s)/participant(s) | J.Welles, R. Miebach |
|---|---|
| Contact for lead author | Chico Soil Survey Office, Chico, CA |
| Date | 09/25/2013 |
| Approved by | |
| Approval date | |
| Composition (Indicators 10 and 12) based on | Annual Production |
Indicators
-
Number and extent of rills:
Rills were noted on steep slopes following heavy precipitation events. Approximate number and distance 2-4 per 100 feet. -
Presence of water flow patterns:
Water flow patterns are generally short flows of 200-500 feet. -
Number and height of erosional pedestals or terracettes:
Some minor pedestals or terracettes might be found on this site, though uncommon. -
Bare ground from Ecological Site Description or other studies (rock, litter, lichen, moss, plant canopy are not bare ground):
Bare ground ranges from 10 to 27 percent. -
Number of gullies and erosion associated with gullies:
No gullies were noted. -
Extent of wind scoured, blowouts and/or depositional areas:
No wind scour or blowouts were noted. -
Amount of litter movement (describe size and distance expected to travel):
Very little if any litter movement was noted. Foothill pine tree litter would be 6 to 8 inches in length by 1/10th of an inch; oak tree litter would be 3 to 4 inches in length by 2 inches wide; annual grass litter would be 4 to 6 inches in length by 1/10 inch wide; forb litter would be 1 to 2 inches in length by 1/4 to 1/2 inch wide. -
Soil surface (top few mm) resistance to erosion (stability values are averages - most sites will show a range of values):
Soils generally have a surface layer that is heavy clay loam or light clay. Permeability is slow, runoff rapid and erosion hazard is severe. -
Soil surface structure and SOM content (include type of structure and A-horizon color and thickness):
0 to 5 inches; brown (7.5YR 5/4) clay loam, dark brown (7.5YR 3/4) moist; moderate coarse granular structure; hard, friable, sticky, plastic; many fine roots; about 12 percent coarse fragments; neutral (pH 6.9); abrupt wavy boundary. (4 to 10 inches thick). 0.5 to 3.0 percent SOM -
Effect of community phase composition (relative proportion of different functional groups) and spatial distribution on infiltration and runoff:
Annual Grasses 60%
Trees 25%
Annual Forbs 10%
Shrubs 5%
High grass cover on footslopes should prevent soil loss from rainfall impact. Backslopes would have less grass cover and more tree cover. The presence of trees and shrubs aids water infiltration and grass cover slows runoff. Trees and low to moderate shrub cover intercept rainfall slowing infiltration. Soil cover of grasses and forbs decreases runoff. -
Presence and thickness of compaction layer (usually none; describe soil profile features which may be mistaken for compaction on this site):
None noted. -
Functional/Structural Groups (list in order of descending dominance by above-ground annual-production or live foliar cover using symbols: >>, >, = to indicate much greater than, greater than, and equal to):
Dominant:
Annual Grasses>Sub-dominant:
Trees>>Other:
Shrubs = Forbs
Additional:
-
Amount of plant mortality and decadence (include which functional groups are expected to show mortality or decadence):
Plant mortality highest in annual grasses and forbs after May through June drought conditions. Foothill pine may show up to 20 percent mortality and oak 1-2 percent mortality -
Average percent litter cover (%) and depth ( in):
-
Expected annual annual-production (this is TOTAL above-ground annual-production, not just forage annual-production):
Annual production varies. Double sampling production values were collected in a 50-80 percent of normal precipitation year. Unfavorable, normal and favorable production ranges from 400 to 1300 pounds per acre respectively. -
Potential invasive (including noxious) species (native and non-native). List species which BOTH characterize degraded states and have the potential to become a dominant or co-dominant species on the ecological site if their future establishment and growth is not actively controlled by management interventions. Species that become dominant for only one to several years (e.g., short-term response to drought or wildfire) are not invasive plants. Note that unlike other indicators, we are describing what is NOT expected in the reference state for the ecological site:
Invasives such as medusahead and yellow star-thistle do not have the potential to become dominant on this site. -
Perennial plant reproductive capability:
Minor amounts of native perennial grasses exist on the site including Chinook brome amd California Melic. Typically the native perennial grasses face strong competition from non-native grasses and forbs. Wet years with fall and winter rains tend to favor non-native grasses on well drained deep soils (Stromberg et al., 2007).
Print Options
Sections
Font
Other
The Ecosystem Dynamics Interpretive Tool is an information system framework developed by the USDA-ARS Jornada Experimental Range, USDA Natural Resources Conservation Service, and New Mexico State University.
Click on box and path labels to scroll to the respective text.