Natural Resources
Conservation Service
Ecological site F022AC005CA
Cryic Sheltered, Moist Sandy Mountain Slopes
Accessed: 04/07/2026
General information
Approved. An approved ecological site description has undergone quality control and quality assurance review. It contains a working state and transition model, enough information to identify the ecological site, and full documentation for all ecosystem states contained in the state and transition model.
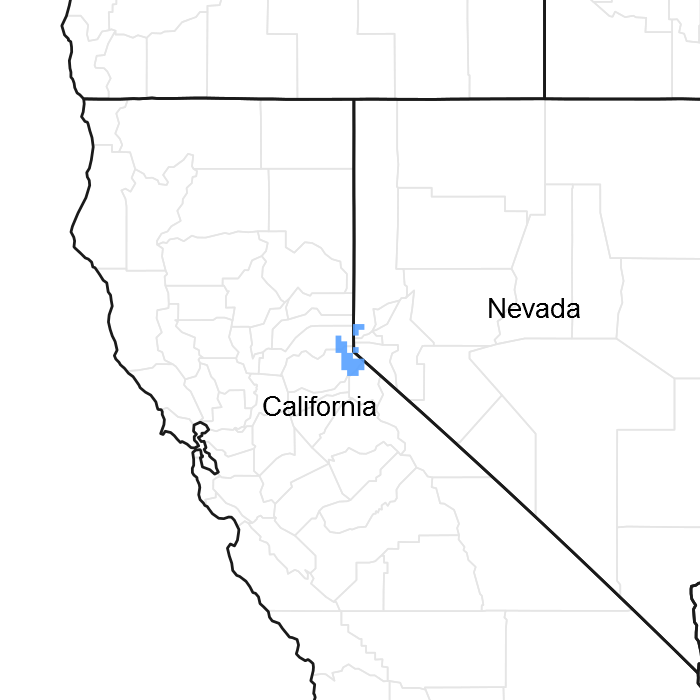
Figure 1. Mapped extent
Areas shown in blue indicate the maximum mapped extent of this ecological site. Other ecological sites likely occur within the highlighted areas. It is also possible for this ecological site to occur outside of highlighted areas if detailed soil survey has not been completed or recently updated.
MLRA notes
Major Land Resource Area (MLRA): 022A–Sierra Nevada and Tehachapi Mountains
MLRA 22A
Major Land Resource Area 22A, Sierra Nevada Mountains, is located predominantly in California and a small section of western Nevada. The area lies completely within the Sierra Nevada Section of the Cascade-Sierra Mountains Province. The Sierra Nevada range has a gentle western slope, and a very abrupt eastern slope. The Sierra Nevada consists of hilly to steep mountains and occasional flatter mountain valleys. Elevation ranges between 1,500 and 9,000 ft throughout most of the range, but peaks often exceed 12,000 ft. The highest point in the continental US occurs in this MLRA (Mount Whitney, 14,494 ft). Most of the Sierra Nevada is dominated by granitic rock of the Mesozoic age, known as the Sierra Nevada Batholith. The northern half is flanked on the west by a metamorphic belt, which consists of highly metamorphosed sedimentary and volcanic rocks. Additionally, glacial activity of the Pleistocene has played a major role in shaping Sierra Nevada features, including cirques, arêtes, and glacial deposits and moraines. Average annual precipitation ranges from 20 to 80 inches in most of the area, with increases along elevational and south-north gradients. Soil temperature regime ranges from mesic, frigid, and cryic. Due to the extreme elevational range found within this MLRA, Land Resource Units (LRUs) were designated to group the MLRA into similar land units.
LRU "C" Northern Sierra Subalpine: Elevations are typically between 7,800 and 9,800 feet. The frost free period is between 30 and 90 days, MAAT is between 35 and 44 degrees, MAP is between 45 and 65 inches. Soils are typically cryic, but frigid soils may occur at lower elevations on southern aspects. Forests are dominated by whitebark pine (Pinus albicaulis), Sierra lodgepole pine (Pinus contorta spp. murrayana), mountain hemlock (Tsuga mertensiana) and/or California red fir (Abies magnifica).
Classification relationships
Forest Alliance = Tsuga mertensiana – Mountain hemlock forest; Association = tentatively Tsuga mertensiana-Pinus contorta spp. murrayana-Pinus monticola. (Sawyer, John O., Keeler-Wolf, Todd, and Evens, Julie M. 2009. A Manual of California Vegetation. 2nd ed. California Native Plant Society Press. Sacramento, California.)
Ecological site concept
This site occurs on deep sandy skeletal soils, over decomposed granite on north facing slopes and cool, moist basins. It typically occurs at elevations between 7800 and 9000 feet, where snow persists late into the season, and the growing season is short. Slopes range from 15 to 70 percent. The vegetation is mixed subalpine forest, with mountain hemlock (Tsuga mertensiana), lodgepole pine (Pinus contorta var. murrayana), red fir (Abies magnifica), and western white pine (Pinus monticola). Understory cover and composition varies with the degree of moisture available at a given location. Purple mountainheath (Phyllodoce breweri) and rose meadowswet (Spiraea splendens) are often abundant, especially in wetter locations.
Associated sites
| F022AC001CA |
Cryic Sandy Mountain Slopes Occurs on adjacent slopes with shallow to moderately deep, sandy skeletal soils at or near treeline. The vegetation is a whitebark pine (Pinus albicaulis) forest with very little understory |
|---|---|
| F022AC003CA |
Frigid-Cryic Sandy Slopes Occurs on adjacent lower elevation sandy slopes. The vegetation is a red-fir (Abies magnifica)- western white pine (Pinus monticola) forest with pinemat mananita (Arctostaphylos nevadensis) dominant in openings. |
| R022AE202CA |
Granitic Pocket Occurs on adjacent lower elevation slopes with a high percentage of rock outcrop and shallow, sandy soils that occur in pockets among bedrock ledges. The vegetation is sparse western juniper (Juniperus grandis)-Jeffrey pine (Pinus jeffreyi)-Sierra lodgepole pine (Pinus contorta var. murrayana) with a sedge-forb community in soil pockets and a huckleberry oak (Quercus vaccinifolia) dominated shrub community among talus. |
| R022AE213CA |
Steep Rubbly Slope Occurs on lower elevation very steep, rubbly slopes with deep soils. The vegetation is a dense shrubland dominated by huckleberry oak (Quercus vaccinifolia) and greenleaf manzanita (Arctostaphylos patula). |
| R022AX101CA |
Frigid Anastomosed System Occurs on adjacent very deep, moist, sandy soils formed in alluvium. This site is dominated by aspen (Populus tremuloides) with a diverse herbaceous understory. |
Similar sites
| F022AC007CA |
North-Facing Cryic Loamy Mountain Slopes This site occurs on moderately deep, andic soils, on north facing slopes. The overstory vegetation is similar, but purple mountainheath and rose meadowsweet are not dominant in the understory. |
|---|---|
| F022AC002CA |
Cryic Sandy North Apsect Mountain Slopes This site occurs in more exposed positions at higher elevations near treeline. Mountain hemlock (Tsuga mertensiana) dominates with whitebark pine (Pinus albicaulis), and there is very little understory vegetation. |
Table 1. Dominant plant species
| Tree |
(1) Tsuga mertensiana |
|---|---|
| Shrub |
(1) Phyllodoce breweri |
| Herbaceous |
Not specified |
Physiographic features
This ecological site is found on mountain slopes at elevations that may from 6,990 to 9,650 feet, but are typically between 7,800 and 9,000 feet. Slopes may range from 5 to 70 percent, but are typically above 15 percent. Aspects are typically north-facing, but may be variable, and runoff class is very low to low.
Table 2. Representative physiographic features
| Landforms |
(1)
Mountain slope
|
|---|---|
| Flooding frequency | None |
| Ponding frequency | None |
| Elevation | 2,131 – 2,941 m |
| Slope | 5 – 70% |
Climatic features
The average annual precipitation ranges from 39 to 65 inches, mostly in the form of snow in the winter months (November through April). The average annual air temperature ranges from 38 to 43 degrees Fahrenheit. The frost-free (>32F) season is 50 to 125 days, and the freeze-free (>28F) season is 90 to 160 days. There are no representative climate stations for this ecological site.
Maximum and minimum monthly climate data for this ESD were generated using PRISM data (PRISM Climate Group, Oregon State University, http://prism.oregonstate.edu, created 4 Feb 2004.) and the ArcGIS ESD extract tool.
Table 3. Representative climatic features
| Frost-free period (average) | 125 days |
|---|---|
| Freeze-free period (average) | 75 days |
| Precipitation total (average) | 1,321 mm |
Figure 2. Monthly precipitation range
Figure 3. Monthly average minimum and maximum temperature
Figure 4. Annual precipitation pattern
Figure 5. Annual average temperature pattern
Influencing water features
This site is not influenced by wetland or riparian water features.
Soil features
The soils associated with this ecological site are deep to paralithic granitic bedrock, and formed in colluvium and/or till over residuum derived from granodiorite. They are excessively drained with rapid permeability. The soil moisture regime is typic xeric and the soil temperature regime is cryic. Surface rock fragments smaller than 3 inches in diameter are typically not present, and larger fragments average 45 percent. The surface texture is very gravelly loamy coarse sand. Subsurface textures are very gravelly loamy coarse sand, and partially decomposed plant material (Oi horizon). Subsurface rock fragments smaller than 3 inches in diameter range from 31 to 35 percent by volume, and larger fragments range from 15 to 45 percent (for a depth of 0 to 49 inches). The soils correlated to this site include Dagget (Sandy-skeletal, mixed Typic Cryorthents).
This ecological site has been correlated with the following mapunits and soil components in the Tahoe Basin soil survey area (CA693):
Area_sym ; Musym ; MUname ; Compname ; Local_phase ; Comp_pct
CA693 ; 9405 ; Dagget very gravelly loamy coarse sand, moist, 15 to 30 percent slopes, rubbly ; Dagget ; moist ; 80
CA693 ; 9406 ; Dagget very gravelly loamy coarse sand, moist, 30 to 70 percent slopes, rubbly ; Dagget ; moist ; 80
CA693 ; 9407 ; Dagget-Rock outcrop complex, moist, 30 to 70 percent slopes ; Dagget ; moist ; 55
CA693 ; 7502 ; Rock Outcrop-Rockbound complex, 30 to 70 percent slopes ; Dagget ; moist ; 5
CA693 ; 7484 ; Meeks gravelly loamy coarse sand, 5 to 15 percent slopes, extremely bouldery ; Dagget ; moist ; 3
CA693 ; 7485 ; Meeks gravelly loamy coarse sand, 15 to 30 percent slopes, extremenly bouldery ; Dagget ; moist ; 3
CA693 ; 7486 ; Meeks gravelly loamy coarse sand, 30 to 70 percent slopes, extremely bouldery ; Dagget ; moist ; 3
CA693 ; 7523 ; Tallac gravelly coarse sandy loam, 30 to 70 percent slopes, very stony ; Dagget ; moist ; 1
Table 4. Representative soil features
| Parent material |
(1)
Colluvium
–
granodiorite
|
|---|---|
| Surface texture |
(1) Very gravelly loamy coarse sand |
| Family particle size |
(1) Sandy |
| Drainage class | Excessively drained |
| Permeability class | Rapid |
| Soil depth | 99 cm |
| Surface fragment cover <=3" | 0% |
| Surface fragment cover >3" | 45% |
| Available water capacity (0-101.6cm) |
4.57 – 6.86 cm |
| Soil reaction (1:1 water) (0-101.6cm) |
5.6 – 6.5 |
| Subsurface fragment volume <=3" (Depth not specified) |
31 – 35% |
| Subsurface fragment volume >3" (Depth not specified) |
15 – 45% |
Ecological dynamics
This site occurs on deep sandy skeletal soils, over decomposed granite on north facing slopes and cool, moist basins. It occurs at high elevations (approximately 7,000 to 9,600 feet), where snow persists late into the season, and the growing season is short.
The vegetation is mixed subalpine forest, with mountain hemlock (Tsuga mertensiana), lodgepole pine (Pinus contorta var. murrayana), red fir (Abies magnifica), and western white pine (Pinus monticola). Mountain hemlock and lodgepole pine are more strongly dominant in wetter locations, such as depressions, bedrock seeps, or high steep north-facing slopes, while red fir and western white pine become more important in locations with less moisture availability. The understory is also variable with soil moisture availability, but purple mountainheath (Phyllodoce breweri) and rose meadowsweet (Spiraea splendens) are common, especially in wetter locations. Relative to other forest types, these forests occupy the smallest geographic extent within the Lake Tahoe Basin (Taylor et al. 2012).
Ecological/Disturbance Factors
Fire and fire suppression, climate fluctuations and climate warming, and pathogen outbreaks are the primary disturbances impacting this ecological site. The high-elevation, poor access landscape that this site occurs in largely escaped the widespread clear-cutting of the Comstock era in the Lake Tahoe Basin (Elliot-Fisk et al. 1996).
The natural fire return interval (FRI) in this forest is relatively long. A Tahoe Basin study estimated an FRI of 55 years for subalpine mountain hemlock – red fir forests (Taylor et al. 2012). Safford (2014) estimated a mean minimum FRI of 100 years and a mean max FRI of 420 years for subalpine forests in California. Studies from other subalpine mountain hemlock systems have estimated an FRI ranging from 45 to over 700 years (Dickman and Cook 1989, Bekker and Taylor 2010, Mallek et al. 2013). Fire ignitions tend to be fewer and fire severity lower in subalpine forests relative to lower elevations due to lower productivity with lower fuel availability, and a shorter fire season due to cooler temperatures and higher relative humidity, and longer and deeper snowpack (Skinner and Chang 1996, Bekker and Taylor 2010, Mallek et al. 2013). These forests experienced a mixed-severity fire regime, with most fires small and of low severity and more rare large, high severity stand-clearing fires (Bekker and Taylor 2010, Mallek et al. 2013). Successional dynamics following stand-clearing fire in these forests are not well studied. In Oregon, lodgepole pine was found to become dominant after stand-clearing fire in mountain hemlock forests, maintaining dominance for up to 200 years (Dickman and Cook 1989). However a southern Cascades study found that composition in red fir-mountain hemlock forests remained fairly constant after fire (Bekker and Taylor 2010). Slightly lower elevation red fir-western white pine forests in the Lake Tahoe Basin that were logged during the Comstock era are currently dominated by lodgepole pine (Taylor 2007), and it is assumed that lodgepole pine, which is an important component of these forests pre-fire, would initially dominate post-fire phases here.
Fire suppression over the last century has impacted contemporary forests, although since fire return intervals are naturally longer in these forests, the impacts are much slower to develop relative to lower elevation forests (Skinner and Chang 1996, Bekker and Taylor 2010, Mallek et al. 2013). The dominant effect of fire suppression is an increase in forest density, without major changes in forest composition as are seen in other forest types (Bekker and Taylor 2010, Mallek et al. 2013, Safford and Van de Water 2014).
Whether observed changes in forest density in the subalpine are due to fire suppression or due to climate warming are difficult to disentangle. Several studies attribute observed patterns of increased forest density in the subalpine more to climate warming over fire suppression (Dolanc et al. 2012, Mallek et al. 2013, Safford and Van de Water 2014). Between 1929 and 2009, stem density of subalpine forests in the Sierra Nevada increased over 30 percent, 63 percent of which comes from small understory trees (Dolanc et al. 2012). Over the same time period, daily minimum temperatures increased by 1.2 degrees Celsius, and precipitation increased (Dolanc et al. 2012). Areas mapped by the U.S. Forest Service as subalpine forest in the 1930s are currently mapped as red fir (Safford and Van de Water 2014), indicating a potential compositional shift in these forests. Mountain hemlock establishment is positively affected by warm wet climatic periods, with historic mountain hemlock forest expansion occurring during these climatic phases (Taylor 1995). Lodgepole pine recruitment is also positively impacted by warm wet periods with low snowpack (Pierce and Taylor 2011). Increases in subalpine forest density, likely due to a warmer climate, are already occurring. Recent California based climate models predict a 9 degree F increase in temperature by 2100, and more conservative models predict a 2 to 4 degree F increase in winter and 4 to 8 degree increase in summer (Safford et al. 2012). Models are more variable for precipitation, but recent models for the Sierra Nevada, predict similar to slightly less precipitation. Most models agree that summers will become drier, since more of the precipitation is predicted to come as rain, and snow melt-off will occur earlier in spring (Hayhoe et al. 2004, Safford et al. 2012). These scenarios will bring more extreme changes to subalpine forests. Species requiring cool, moist conditions to dominate, such as mountain hemlock, will likely be pushed upslope, while species with wider ecological amplitude, such as red fir and lodgepole pine, may remain dominant in this site. With extreme warming, lower montane species like Jeffrey pine (Pinus jeffreyi) and white fir (Abies magnifica) will likely move into the elevation zone occupied by this site. How pathogens, both existing and novel, increased fire frequencies with increased tree densities, introduced species, mycorrhizal relationships, and new species interactions will impact dynamics under these warming scenarios is unknown.
Several pathogens may impact the dominant tree species of this ecological site, but none have been observed to initiate community phase shifts or state transitions. A description of these pathogens may be found in the Other information narrative of the Site Interpretations section of this ESD.
The reference state consists of the most successionally advanced community phase (numbered 1.1) as well as other community phases that result from natural and human disturbances. Community phase 1.1 is deemed the phase representative of the most successionally advanced pre-European plant/animal community including periodic natural surface fires that influenced its composition and production.
All tabular data listed for a specific community phase within this ecological site description represent a summary of one or more field data collection plots taken in communities within the community phase. Although such data are valuable in understanding the phase (kinds and amounts of ground and surface materials, canopy characteristics, community phase overstory and understory species, production and composition, and growth), it typically does not represent the absolute range of characteristics nor an exhaustive listing of species for all the dynamic communities within each specific community phase.
State and transition model
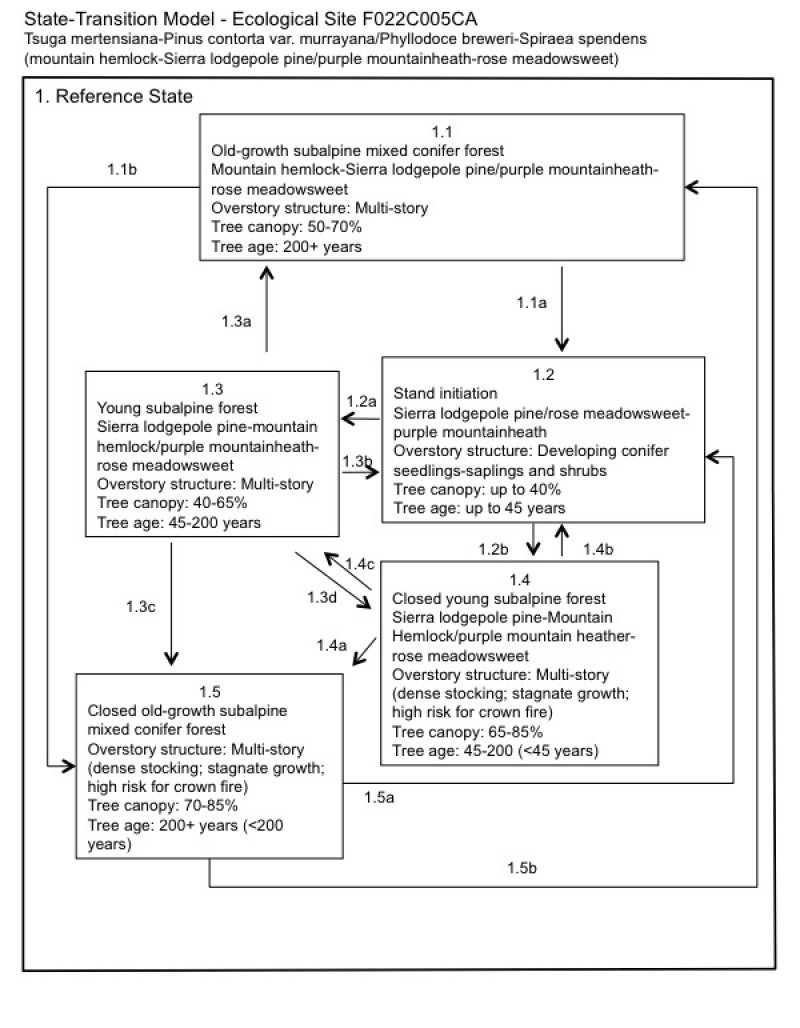
Figure 6. F022AC005CA
More interactive model formats are also available.
View Interactive Models
More interactive model formats are also available.
View Interactive Models
Click on state and transition labels to scroll to the respective text
Ecosystem states
State 1 submodel, plant communities
Communities 1, 5 and 2 (additional pathways)
State 1
Reference
Community 1.1
Old-growth subalpine mixed conifer forest

Figure 7. Community Phase 1.1
This old growth subalpine mixed conifer community phase is fairly undisturbed by human influences because of its inaccessibility. This community phase is maintained by a mixed severity fire regime, dominated by patchy low severity, small fires, often not extending beyond a few trees. Fire suppression has had fairly minimal impacts on many representations of this community, since the duration that fire suppression policies have been in effect is not far out of the range of the natural fire return interval. Thus, this phase is very similar to the historic most successionally advanced community phase. Mountain hemlock (Tsuga mertensiana), Sierra lodgepole pine (Pinus contorta var. murrayana), California red fir (Abies magnifica), and western white pine (Pinus monticola) create a relatively dense multi-layered forest. The trees in this forest are commonly over 200 years old, but are slow growing due to harsh environmental variables, such as a short growing season and low soil fertility. An estimate on age for this community phase ranges from 200 to more than 500 years.
Forest overstory. Mountain hemlock, Sierra lodgepole pine, California red fir, and western white pine provide approximately 50 to 70 percent cover. The forest has several canopy layers, with the height of the upper canopy ranging from 50 to 120 feet, with an average of 65 feet. Tree diameters vary by species, but average 30 inches, with Sierra lodgepole pine and mountain hemlock generally being smaller than California red fir and western white pine.
Forest understory. The cover and composition of the understory varies with site moisture availability. Shrub cover averages 35 percent, but may range from 1 to 70 percent. Purple mountainheath is the dominant shrub, and may form dense patches. Rose meadowsweet may also be abundant. Other shrubs indicative of wet habitats are typically present at lower cover, including dwarf bilberry (Vaccinium cespitosum), western labrador tea (Ledum glandulosum), western moss heather (Cassiope mertensiana), and Sierra willow (Salix orestera). Pinemat manzanita (Arctostaphylos nevadensis) is more abundant in drier locations. Forb cover is low, with an average of 3 percent. Species indicative of wet habitats such as California false hellebore (Veratrum californicum var. californicum), rough Canada goldenrod (Solidago canadensis), high mountain cinquefoil (Potentilla flabellifolia), and tundra aster (Oreostemma alpigenum) are often present, as are species more typical of shady Sierra Nevada forests such as whiteveined wintergreen (Pyrola picta), sidebells wintergreen (Orthilia secunda). A range of other forb species may be present at a particular site. Grass and grasslike cover is also low, averaging 2 percent. Species most frequently present include Parry’s rush (Juncus parryi), pullup muhly (Muhlenbergia filiformis), and shorthair reedgrass (Calamogrostis breweri).
Table 5. Soil surface cover
| Tree basal cover | 2-4% |
|---|---|
| Shrub/vine/liana basal cover | 1-3% |
| Grass/grasslike basal cover | 0% |
| Forb basal cover | 0% |
| Non-vascular plants | 0% |
| Biological crusts | 0% |
| Litter | 30-85% |
| Surface fragments >0.25" and <=3" | 0-15% |
| Surface fragments >3" | 4-25% |
| Bedrock | 0-3% |
| Water | 0% |
| Bare ground | 0-15% |
Table 6. Woody ground cover
| Downed wood, fine-small (<0.40" diameter; 1-hour fuels) | 0-15% |
|---|---|
| Downed wood, fine-medium (0.40-0.99" diameter; 10-hour fuels) | 0-15% |
| Downed wood, fine-large (1.00-2.99" diameter; 100-hour fuels) | 0-15% |
| Downed wood, coarse-small (3.00-8.99" diameter; 1,000-hour fuels) | 0-20% |
| Downed wood, coarse-large (>9.00" diameter; 10,000-hour fuels) | 0-15% |
| Tree snags** (hard***) | – |
| Tree snags** (soft***) | – |
| Tree snag count** (hard***) | |
| Tree snag count** (hard***) |
* Decomposition Classes: N - no or little integration with the soil surface; I - partial to nearly full integration with the soil surface.
** >10.16cm diameter at 1.3716m above ground and >1.8288m height--if less diameter OR height use applicable down wood type; for pinyon and juniper, use 0.3048m above ground.
*** Hard - tree is dead with most or all of bark intact; Soft - most of bark has sloughed off.
Table 7. Canopy structure (% cover)
| Height Above Ground (m) | Tree | Shrub/Vine | Grass/ Grasslike |
Forb |
|---|---|---|---|---|
| <0.15 | 0-1% | 0-10% | 0-2% | 0-1% |
| >0.15 <= 0.3 | 0-5% | 1-50% | 0-4% | 2-7% |
| >0.3 <= 0.6 | 1-9% | 1-60% | 0-2% | 0-2% |
| >0.6 <= 1.4 | 1-9% | 0-45% | – | – |
| >1.4 <= 4 | 1-9% | 0-4% | – | – |
| >4 <= 12 | 5-20% | – | – | – |
| >12 <= 24 | 35-60% | – | – | – |
| >24 <= 37 | 0-55% | – | – | – |
| >37 | – | – | – | – |
Community 1.2
Stand initiation
This community phase develops after severe canopy fire or clear-cut. Detailed data on post-fire succession is not available for the geographic area of this ecological site. Shrubs are likely to dominate the immediate post-fire community. Purple mountainheath may resprout and re-colonize from off-site dispersal after fire (Sawyer et al. 2009). Rose meadowsweet may resprout from rhizomes (Esser 1995), as can dwarf bilberry (Tirmenstein 1990). Dwarf bilberry may become much more abundant in the cleared community phase, forming extensive patches (Agee and Smith 1984, Tirmenstein 1990). Scouler’s willow (Salix scouleriana) responds favorably to disturbance, and may become abundant in this phase (Anderson 2001). Herbaceous species will also likely be more abundant in the immediate post-fire phase, as they are released from the often dense canopy cover of these forests. Canada goldenrod may become abundant after fire (Coladonato 1993). Species such as long-spur lupine (Lupinus arbustus), lambstongue ragwort (Senecio integerrimus), Sierra beardtongue (Penstemon heterodoxus), and mountain pride (Penstemon newberryi) will likely increase, while more shade-loving species like whiteveined and sidebells wintergreen will decline. Remnant overstory trees may be present in limited numbers in this phase, and succession will occur more rapidly if more mature trees remain (Agee and Smith 1984, Pierce and Taylor 2011). Sierra lodgepole pine establishes best in full sun in bare mineral soil (Cope 1993, Pierce and Taylor 2011), and will establish in the spring following fire, possibly dominating the post-fire community phase for up to 200 years (Dickman and Cook 1989). In the Tahoe Basin, Sierra lodgepole pine became and remains dominant in red fir forests after clear-cutting (Taylor 2007). Western white pine is a fire-dependent species that also requires open conditions for regeneration (Griffith 1992), and will establish early post-burn. Since western white pine is not as abundant in this ecological site, or as fast growing as Sierra lodgepole pine, it does not become dominant. Red fir seeds germinate best in bare mineral soil, but seedlings develop best under shade (Cope 1993), so the growth of red fir in this community phase occurs more slowly, after other vegetative cover is established. Mountain hemlock seeds may germinate on bare mineral soil, litter, or even snow if sufficient moisture is available, and seedlings grow best in partial shade (Tesky 1992). Like red fir, the growth of mountain hemlock is slow in this community phase, and it may be centuries before mountain hemlock regains dominance (Agee and Smith 1984, Dickman and Cook 1989).
Community 1.3
Young subalpine forest
Sierra lodgepole pine is dominant in the overstory during with scattered western white pine in drier locations. There is a diverse understory of shrubs and forbs. The growth of shade tolerant conifers, such as California red fir and mountain hemlock increases in the understory of the pines during this phase. This forest community phase develops with occasional canopy disturbances that create a multi-tiered canopy with small gap openings. The natural fire regime consists of small, infrequent, low to moderately intense fires, which are initiated by lightning strikes. These fires may burn understory vegetation or patches of overstory trees. Pest outbreaks and avalanches may also create canopy openings in this forest community phase.
Community 1.4
Closed young subalpine forest
This community phase is defined by a dense canopy and high basal area of mixed subalpine conifers. Sierra lodgepole pine maintains dominance during this phase, but mountain hemlock, which is slower growing and shade tolerant, continues to increase in cover in the understory. In drier positions, California red fir establishes also increases in the understory. This community phase develops more rapidly during periods of warm, wet weather, which increases establishment of understory trees. Canopy cover ranges from 65 to 85 percent. The trees are overcrowded and often diseased and stressed due to the competition for water and nutrients. This stress makes the trees more susceptible to death from disease and drought.
Community 1.5
Closed old-growth subalpine mixed conifer forest
The closed old-growth subalpine mixed conifer forest develops with the continued exclusion of fire and other natural disturbances, allowing tree density to increase. This community phase develops more rapidly during periods of warm, wet weather, which increases establishment of understory trees. Mountain hemlock regains dominance over Sierra lodgepole pine. The estimated age for this community phase ranges from 200 to 500+ years.
Pathway 1.1a
Community 1.1 to 1.2
In the event of a severe canopy fire or clear-cut, the old growth forest would transition to stand initiation, community phase 1.2. Because of the natural fragmentation of this ecological site, low fuels, and short fire season, crown fires are uncommon.
Pathway 1.1b
Community 1.1 to 1.5
If periodic surface fires continue to be excluded from this system for several centuries, tree density may increase, and tree health and vigor may decline. This would shift the community phase towards the old-growth, closed subalpine mixed conifer forest (community phase 1.5). This dense phase may be reached more quickly with sustained warm, wet periods.
Pathway 1.2a
Community 1.2 to 1.3
The natural pathway is to community phase 1.3, a young, open subalpine mixed conifer forest. This pathway is facilitated with a natural fire regime, small pest outbreaks, avalanches, or other natural disturbances that create forest openings.
Pathway 1.2b
Community 1.2 to 1.4
An alternate pathway is created when fire or other natural disturbances do not create openings in the forest structure or canopy. In this case a young, closed subalpine mixed conifer forest develops (community phase 1.4). This pathway is accelerated with warm, wet climatic periods.
Pathway 1.3a
Community 1.3 to 1.1
This is the natural pathway for this community phase, which requires small scale canopy disturbances, such as lighting-ignited surface fires, partial tree mortality from a pest outbreaks, or from avalanches and wind throw. This pathway leads to community phase 1.1.
Pathway 1.3b
Community 1.3 to 1.2
In the event of a canopy fire, this community phase would return to community phase 1.2.
Pathway 1.3d
Community 1.3 to 1.4
If natural disturbances do not occur, then the density of the forest increases. The increased density shifts this community phase (if younger than 200 years) towards the younger closed, subalpine mixed conifer forest, community phase 1.4. This pathway is accelerated with warm, wet climatic periods.
Pathway 1.3c
Community 1.3 to 1.5
If natural disturbances do not occur, then the density of the forest increases. The increased density shifts this community phase (if near or older than 200 years) towards the old-growth subalpine mixed conifer forest, community phase 1.5.
Pathway 1.4b
Community 1.4 to 1.2
A severe canopy fire would return this community to the stand initiation phase (community phase 1.2).
Pathway 1.4c
Community 1.4 to 1.3
Manual treatment to thin out the understory trees and fuels and/or prescribed burns could be implemented to shift this forest back to the relatively open subalpine mixed conifer forest (community phase 1.3). A partial mortality disease or pest infestation could also create a shift towards community phase 1.3.
Pathway 1.4a
Community 1.4 to 1.5
If fire continues to be excluded from this system the old-growth closed subalpine mixed conifer forest, community phase 1.5, develops.
Pathway 1.5b
Community 1.5 to 1.1
Manual treatment to thin out the understory trees and fuels and/or prescribed burns could be implemented to shift this forest back to a relatively open subalpine mixed conifer community (community phase 1.1). A partial mortality disease or pest infestation could also create a shift towards community phase 1.1.
Pathway 1.5a
Community 1.5 to 1.2
A severe canopy fire would initiate a return to the stand initiation phase (community phase 1.2).
Additional community tables
Table 8. Community 1.1 forest overstory composition
| Common name | Symbol | Scientific name | Nativity | Height (m) | Canopy cover (%) | Diameter (cm) | Basal area (square m/hectare) |
|---|---|---|---|---|---|---|---|
|
Tree
|
|||||||
| Sierra lodgepole pine | PICOM | Pinus contorta var. murrayana | Native | – | 1–30 | 10.2–78.7 | – |
| mountain hemlock | TSME | Tsuga mertensiana | Native | – | 9–30 | 10.2–101.6 | – |
| western white pine | PIMO3 | Pinus monticola | Native | – | 0–22 | 10.2–119.4 | – |
| California red fir | ABMA | Abies magnifica | Native | – | 0–10 | 10.2–86.4 | – |
Table 9. Community 1.1 forest understory composition
| Common name | Symbol | Scientific name | Nativity | Height (m) | Canopy cover (%) | |
|---|---|---|---|---|---|---|
|
Grass/grass-like (Graminoids)
|
||||||
| shorthair reedgrass | CABR | Calamagrostis breweri | Native | – | 0–2 | |
| Parry's rush | JUPA | Juncus parryi | Native | – | 0–1 | |
| pullup muhly | MUFI2 | Muhlenbergia filiformis | Native | – | 0–0.5 | |
|
Forb/Herb
|
||||||
| Canada goldenrod | SOCA6 | Solidago canadensis | Native | – | 0.5–1 | |
| mountain pride | PENE3 | Penstemon newberryi | Native | – | 0–1 | |
| Sierra beardtongue | PEHE2 | Penstemon heterodoxus | Native | – | 0–1 | |
| California false hellebore | VECAC2 | Veratrum californicum var. californicum | Native | – | 0–1 | |
| longspur lupine | LUAR6 | Lupinus arbustus | Native | – | 0–1 | |
| sidebells wintergreen | ORSE | Orthilia secunda | Native | – | 0–0.5 | |
| tundra aster | ORALA3 | Oreostemma alpigenum var. andersonii | Native | – | 0–0.5 | |
| lambstongue ragwort | SEIN2 | Senecio integerrimus | Native | – | 0–0.5 | |
| whiteveined wintergreen | PYPI2 | Pyrola picta | Native | – | 0–0.5 | |
| high mountain cinquefoil | POFL3 | Potentilla flabellifolia | Native | – | 0–0.5 | |
|
Shrub/Subshrub
|
||||||
| purple mountainheath | PHBR4 | Phyllodoce breweri | Native | – | 0–60 | |
| rose meadowsweet | SPSPS | Spiraea splendens var. splendens | Native | – | 0–30 | |
| western moss heather | CAME7 | Cassiope mertensiana | Native | – | 0–6 | |
| dwarf bilberry | VACE | Vaccinium cespitosum | Native | – | 0–3 | |
| gooseberry currant | RIMO2 | Ribes montigenum | Native | – | 0–3 | |
| sticky currant | RIVI3 | Ribes viscosissimum | Native | – | 0–3 | |
| pinemat manzanita | ARNE | Arctostaphylos nevadensis | Native | – | 0–3 | |
| Scouler's willow | SASC | Salix scouleriana | Native | – | 0–2 | |
| Sierra willow | SAOR | Salix orestera | Native | – | 0–2 | |
| western Labrador tea | LEGL | Ledum glandulosum | Native | – | 0–1 | |
|
Tree
|
||||||
| Sierra lodgepole pine | PICOM | Pinus contorta var. murrayana | Native | – | 1–3 | |
| mountain hemlock | TSME | Tsuga mertensiana | Native | – | 1–3 | |
| California red fir | ABMA | Abies magnifica | Native | – | 0.5–1 | |
Interpretations
Animal community
Almost 50 bird species and many mammals use this forest for food, cover, or habitat. Common animals include bears, deer, chipmunks, and squirrels. Dead or dying trees provide nesting sites for cavity-nesting birds, and the fallen branches from these trees provide sites for ground-nesting birds and mammals. Conifr seeds are a food source for squirrels, chipmunks, birds, and mice (Cope 1993, Tesky 1992).
Recreational uses
This area provides beautiful scenery and is used for backpacking, hiking, bike riding, photography, and other activities.
Wood products
The remoteness of this area and the slow growth of trees at this elevation make commercial forestry production impractical. However, the wood of Sierra lodgepole pine, mountain hemlock, California red fir and western white pine is of good quality. Sierra lodgepole pine is suited for common lumber grades, and used for light framing materials, interior paneling, exterior trim, posts, railroad ties, pulp and paper, and has potential for structural particle board. The uniform size of Sierra lodgepole pine makes harvesting efficient (Cope 1993).
The wood of California red fir is of high quality and is stronger than other firs. It is used for fuel, coarse lumber, quality veneers, solid framing, plywood, printing paper, and high-quality wrapping paper, and is preferred for pulping (Cope 1993).
Western white pine can produce valuable timber and is often used for finish work. It is used to build doors, paneling, dimension stock, matches, and toothpicks. The wood is also excellent for carving (Griffith 1992).
The wood of mountain hemlock is moderately strong and light colored. It is most often used for small-dimension lumber and pulp. The wood is also used for railway ties, mine timbers, interior finish, crates, kitchen cabinets, and flooring and ceilings (Tesky 1992).
Other information
Site index documentation:
Schumacher (1928), Alexander (1966), Barnes(1962) and Dunning (1942) were used to determine forest site productivity for red fir, Sierra lodgepole pine, mountain hemlock and western white pine, respectively. Low to High values of Site index and CMAI (culmination of mean annual increment) give an indication of the range of inherent productivity of this ecological site. (CMAI values are not available for western white pine, so zeros were used to indicate the lack of data.) Site index relates to height of dominant trees over a set period of time and CMAI relates to the average annual growth of wood fiber in the boles/trunks of trees. Site index and CMAI listed in the Forest Site Productivity section are in units of feet and cubic feet/acre/year, respectively. Both site index and CMAI are estimates; on-site investigation is recommended for specific forest management units for each soil classified to this ecological site. The historical and actual basal area of trees within a growing stand will greatly influence CMAI.
Trees appropriate for site index measurement typically occur in stands of community phases 1.3 and 1.4. Site trees are selected according to guidance in reference publications. Please refer to the Tahoe Basin Soil Survey for detailed site index information by soil component.
Pathogen information:
Several pathogens can affect mountain hemlock, but its patchy distribution in this area makes epidemics rare. These pathogens include laminated root rot (Phellinus weiri), heart rots, such as Indian paint fungus (Echinodontium tinctorum), several needle diseases, snow mold (Herpotrichia nigra), and dwarf-mistletoe (Arceuthobium tsugense) (Tesky 1992).
Sierra lodgepole pine has been killed by the mountain pine beetle (Dendroctonus ponderosae) at lower elevations, but beetle infestations in this mixed subalpine forest seem to be less severe. Lodgepole pine dwarf mistletoe (Arceuthobium americanum) is present in some areas.
The major pathogens that affect California red fir in this area include: red fir dwarf mistletoe (Arceuthobium abietinum f. sp. magnificae), fir broom rust (Melampsorella caryophyllacearum), annosus root rot (Heterobasidion annosum), and the fir engraver (Scolytus ventralis) (Murphy and Knopp 2000). Other diseases also affecting red fir are known as heart rots, which cause the centers of limbs and trunks to decay. Commonly seen heart rots include: yellow cap fungus (Pholiota limonella) and Indian paint fungus (Echinodontium tinctorium). Common pests affecting red fir are cone maggots (Earomyia spp.), several chalcids (Megastigmus spp.) and cone moths (Barbara spp. and Eucosma spp.) (Burns and Honkala 1990).
Red fir dwarf mistletoe (Arceuthobium abietinum f. sp. magnificae) is a parasitic plant common in the survey area. Visible symptoms include witches brooms, top kill, stem cancers, and swellings. The vegetative shoots of the dwarf mistletoe are also often present from spring to fall. Infestation of the red fir dwarf mistletoe can cause reduced growth and vigor, which weakens the tree and allows other pathogens to infest the tree. The mistletoe cankers create an entry point for other diseases such as heart rots and the cytospora canker (Cytospora abietis) (Burns and Honkala 1990).
Fir broom rust (Melampsorella caryophyllacearum) causes dense witches brooms with stunted yellow needles, and can damage tree growth by reducing crown development. Mortality is less common in mature trees than in the younger regeneration trees. The infected branch sheds its needles in fall leaving a barren dead looking branch. The alternate host for this rust is the chickweeds (Stellaria spp. and Cerastium spp.) (Hagle et al. 2003)
Annosus root rot (Heterobasidion annosum) can affect large acres of fir forest. It slowly decays the roots, the root collar and the stem butt for many years causing structural weaknesses and making the tree vulnerable to windthrow. Annosus root rot can spread from infected roots to healthy roots as well as aerially by infecting freshly cut stumps or other fresh tree wounds. Painting Borax on the freshly cut stumps restricts the entry of the fungus. In all management activities, it is important to reduce damage to the bark because the rot itself does not often kill red fir directly, but it weakens the tree and makes it easier for bark beetles (Scolytus spp), annosus root rot, or dwarf mistletoe to infect the tree (Burns and Honkala 1990).
The fir engraver (Scolytus ventralis) can cause extensive damage to a red fir forest and outbreaks can cause several acres of trees to die. It can reach epidemic levels when the trees are stressed due to annosus root rot, dwarf mistletoe, drought, or fire damage (Burns and Honkala 1990).
The major pathogen affecting western white pine is white pine blister rust (Cronartium ribicola). It is a non-native disease that was introduced from Europe and Asia in the 1920s. The fungus causes cankers on five needle pines that eventually kill most of the infected trees. Visible symptoms are swollen cankers with an abundance of pitch flowing down the branch or stem. The cankers can eventually girdle the tree, killing the portions above. The leaves on the upper portion eventually turn red and fall (Hagle et al. 2003). Pruning cankers off of infected stems has been shown to be beneficial. Some strains of western white pine have shown resistance to the disease.
Other pathogens that can affect western white pine are: white pine needle cast (Lophodermella arcuata), pine needle cast (Lophodermium nitens), Bifusella lineari, butt-rot fungi, red ring rot (Phellinus pini), root disease (Phaeolus schweinitzii), annosus root rot (Heterobasidion annosum), and Armillaria root disease (Armillaria spp). Insects affecting western white pine include: Mountain pine beetle (Dendroctonus ponderosae), emarginate ips (Ips emarginatus), and ips beetle (Ips montanus) (Taylor and Halpern 1991).
Table 10. Representative site productivity
| Common name | Symbol | Site index low | Site index high | CMAI low | CMAI high | Age of CMAI | Site index curve code | Site index curve basis | Citation |
|---|---|---|---|---|---|---|---|---|---|
| California red fir | ABMA | 21 | 21 | 79 | 79 | 140 | 050 | – | |
| mountain hemlock | TSME | 55 | 55 | 45 | 45 | 70 | 990 | – | |
| Sierra lodgepole pine | PICOM | 53 | 53 | 41 | 41 | 90 | 520 | – | |
| western white pine | PIMO3 | 80 | 80 | 0 | 0 | 0 | 605 | – |
Supporting information
Inventory data references
The following NRCS plots represent community phase 1.1. of this ecological site:
rx03046 - Type location
rx03054
rx03059
rtg02h39
Type locality
| Location 1: El Dorado County, CA | |
|---|---|
| UTM zone | N |
| UTM northing | 4313145 |
| UTM easting | 747419 |
| General legal description | Take Hwy 89 to Eagle falls trailhead to upper Velma Lake. Go down the creek from Upper Velma Lake outlet. The plot is on the north side of the creek. |
Other references
Agee, J. K. and L. Smith. 1984. Subalpine tree reestablishment after fire in the Olympic Mountains, Washington. Ecology 65.
Alexander, Robert R. 1966. Site indexes for Lodgepole pine, with corrections for stand density: instructions for field use. USDA, Forest Service. Rocky Mountain Forest and Range Experiment Station Research Paper RM-24.
Anderson, M. D. 2001. Salix scouleriana. Fire Effects Information System [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory.
Barnes, George H. 1962. Yield of even-aged stands of western hemlock. USDA, Forest Service. Pacific Northwest Forest and Range Experiment Station Technical Bulletin 1273.
Bekker, M. F. and A. H. Taylor. 2010. Fire disturbance, forest structure, and stand dynamics in montane forests of the southern Cascades, Thousand Lakes WIlderness, California, USA. Ecoscience 17:59-72.
Burns, R. M. and B. H. Honkala. 1990. Silvics of North America: 1. Conifers; 2. Hardwoods. U.S Department of Agriculture, Forest Service, Washington, DC.
Coladonato, M. 1993. Solidago canadensis.in O. Fire Effects Information System, editor. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory.
Cope, A. B. 1993. Pinus contorta var. murreyana. Fire Effects Information System [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory.
Dickman, A. and S. Cook. 1989. Fire and fungus in a mountain hemlock forest. Canadian Journal of Botany 67:2005-2016.
Dolanc, C. R., J. H. Thorne, and H. D. Safford. 2012. Widespread shifts in the demographic structure of subalpine forests in the Sierra Nevada, California, 1934-2007. Global Ecology and Biogeography 22:246-276.
Dunning, Duncan. 1942. A site classification for the mixed-conifer selection forest of the Sierra Nevada. USDA, Forest Service. California Forest and Range Experiment Station Research Note 28.
Elliot-Fisk, D. L., R. Harris, R. A. Rowntree, T. C. Cahill, R. Kattelmann, P. Rucks, O. K. Davis, R. Lacey, D. A. Sharkey, L. Duan, D. Leisz, S. L. Stephens, C. R. Goldman, S. Lindstrom, D. S. Ziegler, G. E. Gruell, and D. Machida. 1996. Lake Tahoe Case Study. Pages 217-276 Sierra Nevada Ecosystem Project. University of California, Centers for Water and Wildland Resources, Davis, CA.
Esser, L. L. 1995. Spiraea douglasii. Fire Effects Information System [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory.
Griffith, R. S. 1992. Pinus monticola. Fire Effects Information System, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory.
Hagle, S. K., K. E. Gibson, and S. Tunnock. 2003. Field guide to diseases and insect pests of northern and central Rocky Mountain conifers. U.S. Department of Agriculture, Forest Service, State and Private Forestry, Intermountain Region.
Mallek, C., H. Safford, J. Viers, and J. Miller. 2013. Modern departures in fire severity and area vary by forest type, Sierra Nevada and southern Cascades, California, USA. Ecosphere 4:1-28.
Murphy, D. D. and C. M. Knopp. 2000. Lake Tahoe Basin Watershed Assessment. PSW-GTR-175, USDA Forest Service, Pacific Southwest Research Station.
Pierce, A. D. and A. H. Taylor. 2011. Fire severity and seed source influence lodgepole pine (Pinus contorta var. murrayana) regeneration in the southern cascades, Lassen volcanic National Park, California. Landscape Ecology 26:225-237.
Safford, H. D. and K. M. Van de Water. 2014. Using fire return interval departure (FRID) analysis to map spatial and temporal changes in fire frequency on National Forest lands in California. PSW-RP-266, US Department of Agriculture, Forest Service, Pacific Southwest Research Station.
Sawyer, J. O., T. Keeler-Woolf, and J. M. Evans. 2009. A manual of California vegetation. 2nd edition. California Native Plant Society, Sacramento, California.
Skinner, C. N. and C.-R. Chang. 1996. Fire regimes, past and present. Pages 1041-1069 Status of the Sierra Nevada. Sierra Nevada Ecosystems Project: Final report to Congress. University of California, Centers for Water and Wildland Resources, Davis, CA.
Schumacher, Francis X. 1928. Yield, stand and volume tables for red fir in California. University of California Agricultural Experiment Station Bulletin 456.
Taylor, A. H. 1995. Forest expansion and climate change in the mountain hemlock (Tusga mertensiana) zone, Lassen Volcanic National Park, California, U.S.A. Arctic and Alpine Research 27:207-216.
Taylor, A. H. 2007. Forest changes since Euro-American settlement and ecosystem restoration in the Lake Tahoe Basin, USA. US Department of Agriculture, Forest Service, Albany, USA.
Taylor, A. H. and C. B. Halpern. 1991. The structure and dynamics of Abies magnifica forests in the southern Cascade Range, USA. Journal of Vegetation Science 2:189-200.
Taylor, A. H., R. S. Maxwell, C. Skinner, and H. Safford. 2012. Identifying spatially explicit reference conditions for forest landscapes in the LTB, USA.
Tesky, J. L. 1992. Tsuga mertensiana. In: Fire Effects Information System [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory.
Tirmenstein, D. 1990. Vaccinium caespitosum. Fire Effects Information System [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory.
Contributors
Alice Miller
Lyn Townsend
Marchel M. Munnecke
Marchel Munnecke
Rangeland health reference sheet
Interpreting Indicators of Rangeland Health is a qualitative assessment protocol used to determine ecosystem condition based on benchmark characteristics described in the Reference Sheet. A suite of 17 (or more) indicators are typically considered in an assessment. The ecological site(s) representative of an assessment location must be known prior to applying the protocol and must be verified based on soils and climate. Current plant community cannot be used to identify the ecological site.
| Author(s)/participant(s) | |
|---|---|
| Contact for lead author | |
| Date | |
| Approved by | |
| Approval date | |
| Composition (Indicators 10 and 12) based on | Annual Production |
Indicators
-
Number and extent of rills:
-
Presence of water flow patterns:
-
Number and height of erosional pedestals or terracettes:
-
Bare ground from Ecological Site Description or other studies (rock, litter, lichen, moss, plant canopy are not bare ground):
-
Number of gullies and erosion associated with gullies:
-
Extent of wind scoured, blowouts and/or depositional areas:
-
Amount of litter movement (describe size and distance expected to travel):
-
Soil surface (top few mm) resistance to erosion (stability values are averages - most sites will show a range of values):
-
Soil surface structure and SOM content (include type of structure and A-horizon color and thickness):
-
Effect of community phase composition (relative proportion of different functional groups) and spatial distribution on infiltration and runoff:
-
Presence and thickness of compaction layer (usually none; describe soil profile features which may be mistaken for compaction on this site):
-
Functional/Structural Groups (list in order of descending dominance by above-ground annual-production or live foliar cover using symbols: >>, >, = to indicate much greater than, greater than, and equal to):
Dominant:
Sub-dominant:
Other:
Additional:
-
Amount of plant mortality and decadence (include which functional groups are expected to show mortality or decadence):
-
Average percent litter cover (%) and depth ( in):
-
Expected annual annual-production (this is TOTAL above-ground annual-production, not just forage annual-production):
-
Potential invasive (including noxious) species (native and non-native). List species which BOTH characterize degraded states and have the potential to become a dominant or co-dominant species on the ecological site if their future establishment and growth is not actively controlled by management interventions. Species that become dominant for only one to several years (e.g., short-term response to drought or wildfire) are not invasive plants. Note that unlike other indicators, we are describing what is NOT expected in the reference state for the ecological site:
-
Perennial plant reproductive capability:
Print Options
Sections
Font
Other
The Ecosystem Dynamics Interpretive Tool is an information system framework developed by the USDA-ARS Jornada Experimental Range, USDA Natural Resources Conservation Service, and New Mexico State University.
Click on box and path labels to scroll to the respective text.