Natural Resources
Conservation Service
Ecological site F022BI110CA
Frigid Humic Loamy Gentle Slopes
Accessed: 04/06/2026
General information
Provisional. A provisional ecological site description has undergone quality control and quality assurance review. It contains a working state and transition model and enough information to identify the ecological site.
Figure 1. Mapped extent
Areas shown in blue indicate the maximum mapped extent of this ecological site. Other ecological sites likely occur within the highlighted areas. It is also possible for this ecological site to occur outside of highlighted areas if detailed soil survey has not been completed or recently updated.
MLRA notes
Major Land Resource Area (MLRA): 022B–Southern Cascade Mountains
Site concept:
Landform: (1) Glacial-valley walls, (2) Strath terrace, (3) Outwash terrace
Elevation (feet): 5,256-7,410
Slope (percent): 2-80, but generally 2 to 40 percent
Water Table Depth (inches): n/a
Flooding-Frequency: None
Ponding-Frequency: None
Aspect: South, East, West
Mean annual precipitation (inches): 43.0-91.0
Primary precipitation: Winter months in the form of snow
Mean annual temperature: 41 to 43 degrees F (5 to 6 degrees C)
Restrictive Layer: Dense till, duripan or bedrock
Temperature Regime: Frigid
Moisture Regime: Xeric
Parent Materials: Colluvium, outwash, glacial till, or in alluvium over residuum from volcanic rocks
Surface Texture: (1) Extremely gravelly medial fine sandy loam, (2) gravelly ashy Sandy loam, (3) Gravelly medial sandy loam
Surface Fragments <=3" (% Cover): 29-85
Surface Fragments > 3" (% Cover): 0-10
Soil Depth (inches): 20-60+
Vegetation: Large mature white fir (Abies concolor), Jeffrey pine (Pinus jeffreyi), incense cedar, (Calocedrus decurrens) and sugar pine (Pinus lambertiana). The reference community phase is similar to the historic community phase but has become denser since the practice of fire suppression. The understory contains a sparse cover of forbs, graminoids and shrubs.
Notes: This white fir-mixed conifer community phase is found at the lowest elevations in the southeast corner of Lassen Volcanic National Park.
Classification relationships
Forest Alliance = Abies concolor – White fir forest; Association = Abies concolor-Calocedrus decurrens-Pinus jeffreyi. (Sawyer, John O., Keeler-Wolf, Todd, and Evens, Julie M. 2009. A Manual of California Vegetation. 2nd ed. California Native Plant Society Press. Sacramento, California.)
Associated sites
| F022BI120CA |
Frigid Gravelly Sandy Loam Outwash-Stream Terraces This is a white fir-Sierra lodgepole pine forest found in slightly wetter conditions. |
|---|---|
| F022BI121CA |
Frigid Shallow Or Moderately Deep Medial-Skeletal Slopes This is an open Jeffrey pine forest with a shrubby understory found on ridges and shoulders. |
| R022BI210CA |
Frigid Loamy Flood Plains This is a riparian complex associated with Hot Springs Creek. |
| R022BI211CA |
Spring Complex This site is associated with springs, generally dominated by alder. |
| R022BI215CA |
Frigid Gravelly Flood Plains This is a riparian complex associated with Kings Creek. |
Similar sites
| F022BI119CA |
Low Precip Frigid Sandy Moraine Slopes This is a white fir-Jeffrey pine forest, which tends towards Jeffrey pine due to lower precipitation. |
|---|---|
| F022BI103CA |
Frigid Tephra Over Slopes And Flats This is a white fir-Jeffrey pine forest found on the west side of the park, with dense shrub phase. |
Table 1. Dominant plant species
| Tree |
(1) Abies concolor |
|---|---|
| Shrub |
(1) Ceanothus cordulatus |
| Herbaceous |
(1) Achnatherum |
Physiographic features
This site is found on back slopes of Glacial-valley walls, colluvial aprons and landslides, strath terraces, outwash terraces, stream terraces and moraines. This site is generally found between 5,200 and 6,500 feet in elevation but is mapped higher in some areas. This site is on slopes that range from 2 to 40 percent, but is associated with map units with up to 80 percent slopes.
Table 2. Representative physiographic features
| Landforms |
(1)
Strath terrace
(2) Outwash terrace |
|---|---|
| Flooding frequency | None |
| Ponding frequency | None |
| Elevation | 5,256 – 7,410 ft |
| Slope | 2 – 40% |
| Aspect | E, S, W |
Climatic features
This ecological site receives most of its annual precipitation in the winter months in the form of snow. The mean annual precipitation ranges between 43 to 91 inches (1,092 to 2,300 mm) and the mean annual temperature is about 42 degrees F (5.5 degrees C). The frost free (>32 degrees F) season is 70 to 90 days. The freeze free (>28 degrees F) season is 85 to 200 days.
There are no representative climate stations for this site.
Table 3. Representative climatic features
| Frost-free period (average) | 90 days |
|---|---|
| Freeze-free period (average) | 200 days |
| Precipitation total (average) | 91 in |
Influencing water features
This ecological site is not influenced by wetland or riparian water features.
Soil features
This site is associated with the following soil components: Kingsiron; Juniperlake, bouldery; Humic Haploxerands, strath terrace; Humic Haploxerands, colluvium; Humic Haploxerands, stream terrace and Humic Haploxerands, outwash terrace. These soils are moderately deep to very deep, well drained, with very low to low AWC. They formed in colluvium, outwash, glacial till, alluvium or in alluvium over residuum from volcanic rocks. The surface textures are gravelly medial sandy loam, extremely gravelly medial fine sandy loam, or gravelly ashy sandy loam, with coarse subsurface textures. Permeability is generally rapid to very rapid through these soils unless dense till or bedrock is encountered then permeability is slow to moderate or very slow respectively.
This ecological site is associated with the following soil components within the Lassen Volcanic National Park Soil Survey Area (CA789):
Map unit, Component, Percent
126 Kingsiron, 45
127 Humic Haploxerands, strath terrace, 65
127 Humic Haploxerands, colluvium, 5
129 Humic Haploxerands, colluvium, 80
129 Kingsiron, 10
129 Humic Haploxerands, strath terrace, 2
160 Humic Haploxerands, stream terrace, 35
164 Humic Haploxerands, strath terrace, 1
166 Humic Haploxerands, outwash terrace, 40
166 Juniperlake, 4
166 Humic Haploxerands, strath terrace, 3
176 Juniperlake, 85
176 Humic Haploxerands, outwash terrace, 3
Table 4. Representative soil features
| Family particle size |
(1) Sandy |
|---|---|
| Drainage class | Well drained |
| Permeability class | Very rapid to very slow |
| Soil depth | 20 in |
| Surface fragment cover <=3" | 29 – 85% |
| Surface fragment cover >3" | 10% |
| Available water capacity (0-40in) |
0.7 – 3.2 in |
| Soil reaction (1:1 water) (0-40in) |
4 – 6.8 |
| Subsurface fragment volume <=3" (Depth not specified) |
30 – 85% |
| Subsurface fragment volume >3" (Depth not specified) |
40% |
Ecological dynamics
The reference community phase is composed of large mature white fir (Abies concolor), Jeffrey pine (Pinus jeffreyi), incense cedar, (Calocedrus decurrens) and sugar pine (Pinus lambertiana). The reference community phase is similar to the historic community phase but has become denser since the practice of fire suppression. The understory contains a sparse cover of forbs, graminoids and shrubs. This white fir-mixed conifer community phase is found at the lowest elevations in the southeast corner of Lassen Volcanic National Park.
White fir is the most abundant tree in these forests. White fir produces single needles that are 1.2 to 2.8 inches long that are distributed along the young branches. The female cones open and fall apart while still attached to the tree, so cones are not often seen on the forest floor. White fir tends to develop a shallow root system that can graft to other white fir roots and spread root rots (Zouhar, 2001).
Jeffrey pine is commonly co-dominant with white fir. Jeffrey pine produces 3 to 8 inch needles in bundles of three. The female seed cones range from 4.7 to 12 inches in length. Jeffrey pine produces a deep taproot and extensive lateral roots (Gucker, 2007) that are intolerant of wet conditions. Jeffrey pine looks similar to ponderosa pine but has a vanilla like odor in the bark, which is not as yellow. Jeffrey pine is shade intolerant and can be replaced over time by white fir if fire is excluded from the system. Older Jeffrey pines are somewhat adapted to fire because their bark is thick enough to provide protection from moderate intensity fires. Additionally, their branches tend to thin along the lower portion of the tree trunk, leaving the crown 65 to 100 feet above the forest floor.
Incense cedar is a common component in these forests but is never dominant. It produces scale-like leaves that are overlapped and arranged in flattened branches. The cones are small and oblong, ranging from 0.8 to 1.5 inches in length, and hang from the end of the branches (Habeck, 2008). The cones open to 3 parts, somewhat resembling an upside down tulip. Incense cedar is a slower growing species than white fir or Jeffrey pine, but it is shade tolerant and can become dense in the understory in the absence of fire. Young incense cedar is fire intolerant because of its very flammable leaves and fibrous bark. Young trees are usually killed by fire but the thicker bark on older trees provides for more protection (Habeck, 2008). Incense cedar seedlings are slow to develop a deep taproot in the first year and reach about half the length of those comparable for Jeffrey pines, initially permitting the Jeffrey pine to out-compete the incense cedar. After the first year however, incense cedar root lengths are equivalent to Jeffrey pine and other species and eventually develop an extensive spreading root system (Powers and Oliver, 1990).
Sugar pine is not often a dominant tree in these forests, but it is very noticeable because it is frequently the largest in diameter and occasionally an emergent tree above the upper canopy. It has broad sweeping branches with 12 inch long cones dangling from the outer ends. The pine needles are in clusters of 5 and are about 3 inches long. Sugar pine germinates best on mineral soil in sunlight, and seedlings quickly develop a deep tap root. Sugar pine seedlings grow slowly, but growth rates increase after the seedling stage. Sugar pine becomes more shade intolerant with age.
Several factors combine to create a habitat suitable for white fir and Jeffrey pine growth. A study on conifer phenology in the Southern Sierra Nevada describes the environmental factors that affect the initiation and seasonal growth of several species. Jeffrey Pine and white fir are included within this study. Temperature is critical in initiating conifer growth after snowmelt. In the study, trees generally started stem growth about 2 weeks after snow melt, a delay that may be related to the warming of soils and roots. If the snow melt was unusually early, the trees did not begin annual growth until specific air temperatures were reached. It was hypothesized that heavy shrub cover delayed the start of annual growth because the shade kept the soil from warming. The pines in the study began leader growth when the air temperatures reached -4 degrees C (24.8 degrees F), and the firs responded after temperatures reached 2 to 3 degrees C (35.6 to 37.4 degrees F). Pines have heavily insulated terminal buds, whereas the terminal buds of fir trees are less insulated and more susceptible to frost damage. The length of the leader growth is predetermined by growth conditions of the prior year. Primordia of fir needles and pine fascicles are developed the year before leader growth. The internode length between fir needles or pine fascicles is determinate. Therefore leader length is determined by the number of primordia developed. It appears that some conifers will not start leader growth until a specific photoperiod (a ratio of light hours to dark hours during one 24 hour period) is met, even if the snow has melted and the temperatures are warm enough. If drought conditions set in before the leader has reached its determinate length, growth will be terminated prematurely. If precipitation comes after the snow has melted, it can prolong the growing season. Conifer growth ceases with the onset of drought conditions and the decline of water potentials (Royce and Barbour, 2001).
This site receives 55 inches average annual precipitation, mostly in the form of snow in winter. As the snow melts it fills macropores in the soil with water. Soil characteristics such as depth and texture determine how much water the soil can hold and how long it will remain before filtering through, evaporating away, or being lost to evapotranspiration. The soils associated with this site have very low to low water holding capacities. Under the same climatic conditions, drought would come earlier to these soils than those with higher water holding capacities. The trees on this site have a short growing season due to the duration of snow and early drought conditions.
Most of the forest within the present park boundary was never logged, but fire suppression has created a change in the stand structure and composition. Historically, with a natural fire regime, this forest would most likely be dominated by large Jeffrey pine and sugar pine in the overstory with a lesser component of white fir and incense cedar. Low to moderate intensity fires would maintain an open forest with patches of montane shrubs, forbs and grasses in the canopy openings. In the absence of fire, white fir would continue to regenerate in the understory, increasing forest density and fuels. Today the forest is multilayered, dense and shady, dominated by white fir. Vegetation on the forest floor is almost nonexistent.
Fire regime studies, using tree rings and fire scars, report historic median fire return intervals in Jeffrey pine-white fir forests of 14, 18.8, and 70 years (Bekker and Taylor; Skinner and Chang; Taylor and Solem respectively). Beaty and Taylor report that fire frequency and intensity is additionally associated with slope position, aspect, and climatic fluctuations. Fire return intervals are longer for north facing slopes than for south facing slopes, and fire intensity increases from lower slopes to upper slope positions. Their study also indicates a slightly later burn season in the Southern Cascades than in the Sierra Nevada. The fire scars in the Southern Cascade are primarily found at the annual tree ring boundary, signifying that the trees were dormant at the time of the fire, whereas in the Sierra Nevada fires, scars are often found in the late-season wood. This timing shift may be due to the timing of summer drought conditions, which begin earlier in the south. In July and August, thunderstorms are common in Lassen Volcanic National Park and the summer drought conditions begin, initiating the fire season. Large fires and multiple small fires in the same season are associated with dry and very dry years (Beaty and Taylor, 2001). Beaty and Taylor report that stand replacing fire is more common on the upper slopes, while low to moderate intensity fires occur only along the lower slopes. This is probably due to the tendency of fire to burn upslope, preheating the fuels as it goes (Beaty and Taylor, 2001). After a stand replacing fire, evenly aged forests are formed. The current management practice of fire suppression has shifted forest density and composition. Fire suppression creates a change in species composition by allowing the fire-intolerant and shade-tolerant firs to increase in cover and density, eventually out-competing the fire-tolerant and shade-intolerant pines (Tayler and Solem, 2001).
Tree pathogens and insect infestations can have significant impacts on the composition and structure of mid and upper montane coniferous forests. Although small infestations may affect relatively few trees, large outbreaks may kill the dominant trees over large areas of forest, creating large canopy openings and stand regeneration. Most of these pathogens and infestations are natural cycles of regulation and can push the closed forest types to more open forests. Fuel loads are often high after outbreaks, creating ideal conditions for high intensity fires.
Pathogens that affect white fir are the dwarf mistletoe (Arceuthobium abietinum f. sp. concoloris), Cytospora canker (Cytospora abietis), broom rust (Melampsorella caryophyllacearum), annosus root disease (Heterobasidium annosum), trunk rot (Echinodontium tinctorium) and the fir engraver (Scotylus ventralis). The most threatening of these is the combination of the fir engraver and annosus root disease. These pathogens can kill large areas of white fir (Bohne, 2006; Laacke, 1990).
Jeffrey Pine is susceptible to several diseases and insect infestations, especially in periods of drought or when overcrowded. Pathogens that affect Jeffrey pine in this area are the dwarf mistletoe (Arceuthobium campylopodum), root disease (Phaeoleus schweinitzii), needle cast (Elytroderma deformans), Jeffrey pine bark beetle, (Dendroctonus jeffreyi), Red turpentine beetle (D. valens), and pine engravers (Ips species). The most threatening of these are the dwarf mistletoe and the Jeffrey pine bark beetle (Bohne, 2006; Jenkinson, 1990).
The most destructive pathogen for incense cedar is pocket dry rot (Tyromyces amarus). Trees are generally infected when the mycelium is able to enter in areas where the bark has been removed by fire, mechanical injury, or other processes. Root diseases are to blame for the majority of incense cedar mortalities. The major root diseases are annosus root disease (Heterobasidium annosum), Armillaria root disease (Armillaria sp.), and laminated root rot (Phellinus weirii) (Powers and Oliver, 1990).
The major pathogens that affect sugar pine (Pinus lambertiana) are white pine blister rust (Cronartium ribicola) and bark beetles (Dendroctonus ponderosae and Ips species). White pine blister rust is an introduced fungus that causes cankers that girdle and eventually kill the infected tree. This fungus is a serious threat to all 5 needle pines but especially sugar pine. White pine blister rust kills old, young, and seedling trees. It can severely inhibit regeneration in infested areas, almost eliminating sugar pine from those areas. The mountain pine beetle (Dendroctonus ponderosae) can cause widespread mortality as well. Please see the section titled Other Information for further details about the tree pathogens mentioned above.
The reference state consists of the most successionally advanced community phase (numbered 1.1) as well as other community phases which result from natural and human disturbances. Community phase 1.1 is deemed the phase representative of the most successionally advanced pre-European plant/animal community including periodic natural surface fires that influenced its composition and production. Because this phase is determined from the oldest modern day remnant forests and/or historic literature, some speculation is necessarily involved in describing it.
All tabular data listed for a specific community phase within this ecological site description represent a summary of one or more field data collection plots taken in communities within the community phase. Although such data are valuable in understanding the phase (kinds and amounts of ground and surface materials, canopy characteristics, community phase overstory and understory species, production and composition, and growth), it typically does not represent the absolute range of characteristics nor an exhaustive listing of species for all the dynamic communities within each specific community phase.
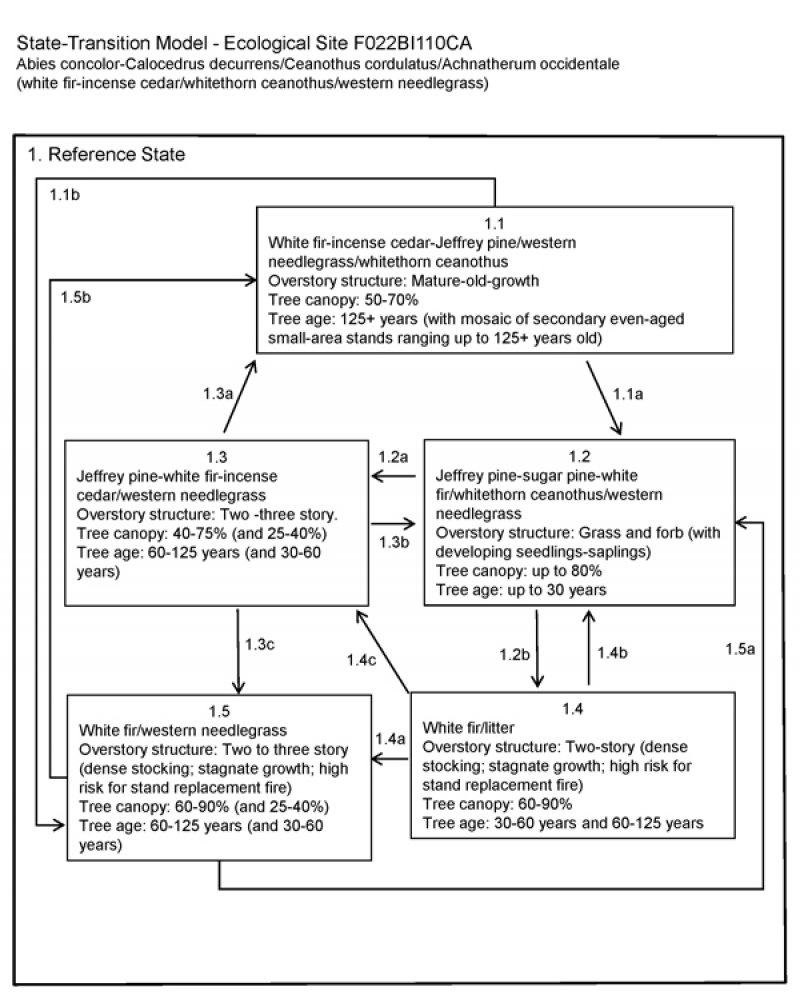
State and transition model
More interactive model formats are also available.
View Interactive Models
More interactive model formats are also available.
View Interactive Models
Click on state and transition labels to scroll to the respective text
Ecosystem states
State 1 submodel, plant communities
State 1
Reference
Community 1.1
White fir-incense cedar-Jeffrey pine/western needlegrass/whitethorn ceanothus
This is the reference community phase for this ecological site, comprising a multi-tiered forest of white fir, Jeffrey pine, sugar pine, and incense cedar. This community phase develops 150 to 300 years after a major disturbance event such as fire. The trees are large with wide reaching canopies. White fir and Jeffrey pine dominate, with several large sugar pines and incense cedars. This forest develops with frequent low intensity fires or occasional small high severity fires that either remove understory trees or create small openings in the forest for gap regeneration. This community phase needs a continued disturbance from low intensity fires to maintain the open understory and reduce competition between the trees for water, nutrients, and sunlight. It is difficult to find a representative site for this community phase because most of the area has missed several fire rotations, resulting in high cover of white fir in the understory. The photo for this community phase is from the type location, which does have several large Jeffrey pine, white fir and sugar pines in the overstory, but the dense understory of white fir is visible. This photo accurately represents a forest on a pathway to the closed white fir forest (Community Phase 1.5).
Forest overstory. Canopy cover ranges from 50 to 70 percent. The structure in this old growth mixed conifer forest is complex, including scattered or clumped trees larger than 30 inches dbh and exceeding 120 feet in height. White fir and Jeffrey pine co-dominate, with a significant portion of sugar pine in the upper canopy. The dominant trees on this site may be 300 to 500 years old. There are two to three understory layers dominated by white fir and incense cedar.
Forest understory. The understory is relatively sparse, having an average of 10 percent cover. Grasses generally dominate with a few forbs and shrubs. Common plants include: needlegrass (Achnatherum spp.), California brome (Bromus carinatus), Orcutt's brome (Bromus orcuttianus), Brainerd's sedge (Carex brainerdii), Ross' sedge (Carex rossii), whitethorn ceanothus (Ceanothus cordulatus), squirreltail (Elymus elymoides), naked buckwheat (Eriogonum nudum), spreading groundsmoke (Gayophytum diffusum) and Sierra gooseberry (Ribes roezlii).
Figure 2. Annual production by plant type (representative values) or group (midpoint values)
Table 5. Annual production by plant type
| Plant type | Low (lb/acre) |
Representative value (lb/acre) |
High (lb/acre) |
|---|---|---|---|
| Shrub/Vine | 0 | 31 | 368 |
| Grass/Grasslike | 2 | 31 | 83 |
| Tree | 1 | 16 | 25 |
| Forb | 0 | 5 | 11 |
| Total | 3 | 83 | 487 |
Table 6. Ground cover
| Tree foliar cover | 50-75% |
|---|---|
| Shrub/vine/liana foliar cover | 0-35% |
| Grass/grasslike foliar cover | 1-40% |
| Forb foliar cover | 0-19% |
| Non-vascular plants | 0% |
| Biological crusts | 0% |
| Litter | 50-80% |
| Surface fragments >0.25" and <=3" | 30-80% |
| Surface fragments >3" | 0-10% |
| Bedrock | 0% |
| Water | 0% |
| Bare ground | 0-2% |
Community 1.2
Jeffrey pine-sugar pine-white fir/whitethorn ceanothus/western needlegrass
This community phase develops when the majority of the overstory trees succumb to a high intensity canopy fire. There may be a few surviving overstory trees, which become an important seed source for regeneration. The mature Jeffrey pine and sugar pines have thicker bark and higher tree branches than white fir and incense cedar and are more likely to survive a fire and supply seed for regeneration. Because Jeffrey pine and sugar pine seedlings germinate well in full sun and mineral soils after fire and white fir and incense cedar prefer partial shade, the pines have an advantage in this early phase of regeneration which assures their existence and prevalence in older stands. Whitethorn ceanothus (Ceanothus cordulatus) is the dominate shrub in this area, and is dependent upon fire for regeneration. It produces abundant seeds which accumulate over the years near the source. The seeds generally remain dormant until fire scarifies the seed coat. Fire scarification followed by a period of cold scarification creates maximum germination rates. It also resprouts from the lignotuber after fire. (Reeves, 2006). Cover from native cool season bunch grasses may be up to 30 percent after a fire. Western needlegrass (Achnatherum occidentale), Columbia needlegrass (Achnatherum nelsonii), squirreltail (Elymus elymoides), California brome (Bromus carinatus), and Orcutt's brome (Bromus orcuttianus) were documented in this area. These grasses are somewhat resistant to fire, because they are generally only top-killed and can regenerate from the root crown after fire. They also regenerate from on or off-site seed. The photo for this community phase is from a prescribed fire that caused mortality in the overstory trees. There is a high cover of grasses after this burn.
Forest overstory. The overstory trees can be completely absent, or a few surviving trees may provide up to 10 percent canopy cover.
Forest understory. The understory is dominated by grasses for a short duration, and the cover of whitethorn ceanothus increases over the first 5 years. Initially young Jeffrey pine and sugar pine seedlings are abundant. There may be a high mortality rate but enough survive to develop a new forest. White fir and incense cedar establish secondarily, in the shade of the young pines. Grass cover may be high including: western needlegrass (Achnatherum occidentale), Columbia needlegrass (Achnatherum nelsonii), California brome (Bromus carinatus), Orcutt's brome (Bromus orcuttianus), and squirreltail (Elymus elymoides).
Community 1.3
Jeffrey pine-white fir-incense cedar/western needlegrass
This forest community phase develops with the natural fire regime, or with manual thinning and prescribed fires. Low to moderate intensity fires clear the understory and remove fuels before they reach hazardous levels, although severe high-intensity canopy fires are also possible. If Jeffrey pine and sugar pine establish during stand initiation they have a fair percentage of cover in the upper canopy. Jeffrey pine and sugar pine have difficulty regenerating and growing well in the understory of the canopy. Their growth and presence is dependent upon fire or other disturbances to maintain an open forest structure with canopy openings.
Community 1.4
White fir/litter
This community phase is defined by a dense canopy and high basal area of white fir. Canopy cover ranges from 65 to 85 percent. The trees are overcrowded and often diseased and stressed due to competition for water and nutrients. This stress makes the trees more susceptible to death from disease and drought. Fire hazard is high in this community phase , a result of the deep accumulation of litter, standing dead and down trees, and the dense multi-layered structure of the forest.
Community 1.5
White fir/western needlegrass
The mature closed white fir forest develops with the continued exclusion of fire, allowing tree density to increase to unhealthy levels. Competition for water and sunlight continue, and tree health and vigor decreases.
Pathway 1.1a
Community 1.1 to 1.2
The primary threat to a white fir–mixed conifer forest is a severe canopy fire. In the event of a severe canopy fire this community phase would return to the regeneration community phase (Community Phase 1.2).
Pathway 1.1b
Community 1.1 to 1.5
If fire is excluded from the old growth community phase, tree density continues to increase and shifts the community phase toward the closed white fir forest (Community Phase 1.5).
Pathway 1.2a
Community 1.2 to 1.3
1.2a. The natural pathway is to community phase 1.3, the Young Open White Fir-Mixed Conifer Forest. This pathway is followed with natural fire regime. Reports vary on the natural fire return interval; this pathway assumes a relatively frequent surface fire interval of 4 to 12 years. Manual thinning with prescribed burns can imitate the natural cycle and lead to the same open community.
Pathway 1.2b
Community 1.2 to 1.4
An alternate pathway is created when fire is excluded from the system and leads to the Young Closed White Fir-Mixed Conifer Forest (Community Phase 1.4).
Pathway 1.3a
Community 1.3 to 1.1
This is the natural pathway for this community phase, which evolved with a historic fire regime of relatively frequent surface and moderate severity fires, and/or partial tree mortality from a pest outbreak. Manual thinning or prescribed burning can be implemented to replace the natural disturbances that keep this forest open. This pathway leads to the reference community phase(Community Phase 1.1).
Pathway 1.3b
Community 1.3 to 1.2
In the event of a canopy fire this community phase would return to Community Phase 1.2, stand initiation.
Pathway 1.3c
Community 1.3 to 1.5
If fire does not occur, forest density increases. This may favor white fir and incense cedar over Jeffrey pine and sugar pine. The increased density shifts this community phase toward the closed white fir forest(Community Phase 1.5).
Pathway 1.4b
Community 1.4 to 1.2
At this point the density of ground fuels and ladder fuels formed in the mid-canopy create conditions for a high intensity canopy fire. A severe fire would initiate stand regeneration (Community Phase 1.2).
Pathway 1.4c
Community 1.4 to 1.3
The natural event of a moderate or surface fire in this forest is unlikely due to the high fuels. Considerable management efforts would be needed to create the open forest conditions that should exist in this forest if it had developed with fire over time. Manual treatments to thin out the white fir and fuels in the understory, and/or prescribed burns, could be implemented to shift this forest back to its natural state of an open white fir-mixed conifer forest (Community Phase 1.3). A partial mortality disease or pest infestation could also create a shift toward Community Phase 1.3.
Pathway 1.4a
Community 1.4 to 1.5
If fire continues to be excluded from this system the mature closed white fir–mixed conifer forest develops (Community Phase 1.5).
Pathway 1.5b
Community 1.5 to 1.1
The natural event of a moderate or surface fire in this forest is unlikely due to the high fuels. Considerable management efforts would be needed to create the open forest conditions that should exist in this forest if it had developed with fire over time. Manual treatments to thin out the understory trees and fuels, and/or prescribed burns, could be implemented to shift this forest back to its natural state of an open white fir-mixed conifer forest (Community Phase 1.1). A partial mortality disease or pest infestation could also create a shift toward Community Phase 1.1, but tree mortality will increase the already high fuel amounts.
Pathway 1.5a
Community 1.5 to 1.2
At this point a severe fire is likely and would initiate stand regeneration (Community Phase 1.2).
Additional community tables
Table 7. Community 1.1 plant community composition
| Group | Common name | Symbol | Scientific name | Annual production (lb/acre) | Foliar cover (%) | |
|---|---|---|---|---|---|---|
|
Tree
|
||||||
| 1 | Tree (understory only) | 1–25 | ||||
| white fir | ABCO | Abies concolor | 1–10 | 1–5 | ||
| Jeffrey pine | PIJE | Pinus jeffreyi | 0–6 | 0–3 | ||
| sugar pine | PILA | Pinus lambertiana | 0–6 | 0–3 | ||
| incense cedar | CADE27 | Calocedrus decurrens | 0–3 | 0–3 | ||
|
Shrub/Vine
|
||||||
| 2 | Shrub | 0–368 | ||||
| huckleberry oak | QUVA | Quercus vacciniifolia | 0–150 | 0–15 | ||
| whitethorn ceanothus | CECO | Ceanothus cordulatus | 0–100 | 0–20 | ||
| Sierra gooseberry | RIRO | Ribes roezlii | 0–100 | 0–10 | ||
| naked buckwheat | ERNU3 | Eriogonum nudum | 0–18 | 0–12 | ||
|
Grass/Grasslike
|
||||||
| 3 | Grass/Grasslike | 2–83 | ||||
| needlegrass | ACHNA | Achnatherum | 2–48 | 1–30 | ||
| California brome | BRCA5 | Bromus carinatus | 0–20 | 0–5 | ||
| squirreltail | ELEL5 | Elymus elymoides | 0–6 | 0–3 | ||
| Ross' sedge | CARO5 | Carex rossii | 0–5 | 0–4 | ||
| Orcutt's brome | BROR2 | Bromus orcuttianus | 0–2 | 0–1 | ||
| Brainerd's sedge | CABR7 | Carex brainerdii | 0–2 | 0–1 | ||
|
Forb
|
||||||
| 4 | Forb | 0–11 | ||||
| white hawkweed | HIAL2 | Hieracium albiflorum | 0–10 | 0–6 | ||
| spreading groundsmoke | GADI2 | Gayophytum diffusum | 0–1 | 0–1 | ||
Table 8. Community 1.1 forest overstory composition
| Common name | Symbol | Scientific name | Nativity | Height (ft) | Canopy cover (%) | Diameter (in) | Basal area (square ft/acre) |
|---|---|---|---|---|---|---|---|
|
Tree
|
|||||||
| white fir | ABCO | Abies concolor | Native | – | 26–40 | – | – |
| Jeffrey pine | PIJE | Pinus jeffreyi | Native | – | 9–15 | – | – |
| sugar pine | PILA | Pinus lambertiana | Native | – | 11–15 | – | – |
| incense cedar | CADE27 | Calocedrus decurrens | Native | – | 4–10 | – | – |
Table 9. Community 1.1 forest understory composition
| Common name | Symbol | Scientific name | Nativity | Height (ft) | Canopy cover (%) | |
|---|---|---|---|---|---|---|
|
Grass/grass-like (Graminoids)
|
||||||
| needlegrass | ACHNA | Achnatherum | Native | – | 1–30 | |
| California brome | BRCA5 | Bromus carinatus | Native | – | 0–5 | |
| Ross' sedge | CARO5 | Carex rossii | Native | – | 0–4 | |
| squirreltail | ELEL5 | Elymus elymoides | Native | – | 0–3 | |
| Orcutt's brome | BROR2 | Bromus orcuttianus | Native | – | 0–1 | |
| Brainerd's sedge | CABR7 | Carex brainerdii | Native | – | 0–1 | |
|
Forb/Herb
|
||||||
| white hawkweed | HIAL2 | Hieracium albiflorum | Native | – | 0–6 | |
| spreading groundsmoke | GADI2 | Gayophytum diffusum | Native | – | 0–1 | |
|
Shrub/Subshrub
|
||||||
| whitethorn ceanothus | CECO | Ceanothus cordulatus | Native | – | 0–20 | |
| huckleberry oak | QUVA | Quercus vacciniifolia | Native | – | 0–15 | |
| naked buckwheat | ERNU3 | Eriogonum nudum | Native | – | 0–12 | |
| Sierra gooseberry | RIRO | Ribes roezlii | Native | – | 0–10 | |
|
Tree
|
||||||
| white fir | ABCO | Abies concolor | Native | – | 1–5 | |
| incense cedar | CADE27 | Calocedrus decurrens | Native | – | 0–3 | |
| Jeffrey pine | PIJE | Pinus jeffreyi | Native | – | 0–3 | |
| sugar pine | PILA | Pinus lambertiana | Native | – | 0–3 | |
Interpretations
Animal community
White fir forests provide browse, cover and nesting sites for a variety of wildlife species. The type and quality of the wildlife habitat varies with the community type. Mature open forests, closed dense white fir forests, young forests and shrub lands provide different habitats and forage for wildlife. Deer and bear can heavily browse young white fir shoots. Porcupines eat the bark of white fir and can kill saplings. Rodents feed on the cambial tissue. Young seedlings and seeds are eaten by animals as well. Douglas squirrels cut and cache white fir cones before the cones are fully mature.
There are about 33 species of mammals commonly present in the white fir forest type in California and, of these, 7 are generally associated with mature forests. About 123 species of birds are found in the white fir forest type of California and southern Oregon, about 50 of which are associated primarily with mature forests. Many of these birds use mature white fir trees and snags for foraging, roosting, nesting and/or breeding. Included are bald eagle, California spotted owl, brown creeper, pileated woodpecker, white-headed woodpecker, and, when near lakes or streams, osprey. Reptiles in white fir forests are represented by 17 species, mostly at lower elevations, 8 of which are associated with mature forests (Zouhar, 2001).
Recreational uses
These areas are suitable for hiking trails and campsites in the flatter areas.
Wood products
White fir wood is used for framing, plywood and, sometimes, pulpwood. The heartwood of white fir decays rapidly if not properly preserved. White fir wood has a low specific gravity and heat production, hence it provides poor firewood compared to other conifers (Zouhar, 2001).
Jeffrey pine wood is used for lumber. No commercial distinction is made between ponderosa pine and Jeffrey pine lumber (Gucker, 2007).
Incense cedar wood is resistant to decay, making it very desirable for exterior use. This wood is used as mud sills, window sashes, sheathing under stucco or brick veneer construction, greenhouse benches, fencing, poles and trellises. It is also widely used for exterior and interior siding. Much of the top quality incense cedar is used in the manufacture of pencils (Habeck, 2008).
Other products
Jeffrey pine seeds are edible. Jeffrey pine sap was used by Native Americans to treat pulmonary disorders and, later, heptane was distilled from the sap and sold to treat pulmonary problems and tuberculosis. Jeffrey pine heptane was also used to develop the octane scale used to rate petroleum used in automobiles (Gucker, 2007).
Other information
Additional information on the common white fir pathogens:
White fir dwarf mistletoe (Arceuthobium abietinum f. sp. concoloris) is a parasitic plant common in the survey area as evident by witches brooms, top kill, stem cankers and swellings. The vegetative shoots of the dwarf mistletoe are often present from spring to fall. A fungus (Cytospora abietis) kills the branches that are infected with dwarf mistletoe. The reduced vigor makes the tree more susceptible to bark beetle and other diseases. The mistletoe cankers, by creating cracks in the bark, create an entry point for other diseases such as heart rots (Burns and Honkala, 1990).
Fir broom rust (Melampsorella caryophyllacearum) is a disease that causes dense witches brooms with stunted yellow needles. The infected branch sheds its needles in fall, leaving a barren dead looking branch. The alternate host for this rust is the chickweeds (Stellaria spp. and Cerastium spp.). This disease can damage tree growth by reducing crown development. Mortality is less common in mature trees than in younger regeneration trees. Secondary infection is possible from heart rots entering through openings in the infected areas (Burns and Honkala, 1990).
Annosus root rot (Heterobasidion annosum) can affect large acres of fir forest. It spreads from infected roots to healthy roots. It slowly decays the roots, the root collar and the stem butt for many years, causing structural weaknesses and making the tree vulnerable to wind throw. Annosus root rot can also be spread aerially, infecting freshly cut stumps or other fresh tree wounds. Painting Borax on the freshly cut stumps restricts the entry of the fungus. In all management activities it is important to reduce damage to the bark. The rot itself does not often kill red fir directly, but it weakens the tree and makes it easier for bark beetles (Scolytus spp.) to infest the tree (Burns and Honkala, 1990).
The fir engraver beetle (Scolytus ventralis) can cause extensive damage to white fir forests. Outbreaks can cause mortality to several acres of trees. It can reach epidemic levels when the trees are stressed due to drought, annosus root rot, dwarf mistletoe, or fire damage.
Additional information on Jeffrey pine pathogens:
Infections from western dwarf mistletoe (Arceuthobium campylopodium) cause witches brooms, reduced growth and tree mortality. Sticky seeds are spread in fall and infest nearby and understory trees. In years of severe drought dwarf mistletoe has induced 60 to 80 percent of the Jeffery pine mortality (Burns and Honkala, 1990).
Jeffrey pine bark beetles (Dendroctonus jeffreyi) are native beetles that can only reproduce in Jeffrey pine. They are a natural cycle in maintaining forest health. They generally attack older weaker trees, but in times of drought or other disturbances such as lightning or fire, epidemic levels can break out and cause extensive damage to the forest. These beetles infest the lower stem and bole of the trees, usually after pine engraver (Ips pini) infestation in the upper portion of the tree. The beetles slowly destroy the cambium, inhibiting the flow of nutrients. A sign of infestation is the changing color of the pine needles from green to yellow or reddish brown, beginning from the top down (Hagle et al., 2003; Smith, 1971).
Additional information about the mountain pine beetle, which can affect sugar pine, as well as other pines:
The mountain pine beetle (Dendroctonus ponderosae) creates pitch tubes on the bark at the point of entry. Boring dust is evident at the base of the tree. These beetles feed on the phloem layer in the inner bark of the tree, eventually girdling the tree. A blue stain fungus is inoculated into the tree by the beetles and reduces the flow of water. These beetles generally infest trees that are weakened by drought or other stresses and usually kill the tree. The engraver beetles (Ips spp.) are a secondary bark beetle, coming in after the mountain pine beetle. They eat the inner bark of the tree and inoculate the blue stain fungus as mentioned above, but the trees have a lower mortality rate. These beetles can be distinguished by their feeding patterns in the wood, and by the shape of the adults.
SITE INDEX DOCUMENTATION:
Schumacher (1926) was used to determine forest site productivity for white fir. Low to High values of Site index and CMAI (culmination of mean annual increment) give an indication of the range of inherent productivity of this ecological site. Site index relates to height of dominant trees over a set period of time and CMAI relates to the average annual growth of wood fiber in the boles/trunks of trees. Site index and CMAI listed in the Forest Site Productivity section are in units of feet and cubic feet/acre/year, respectively. Both site index and CMAI are estimates; on-site investigation is recommended for specific forest management units for each soil classified to this ecological site. The site index for white fir was quite variable. The historical and actual basal area of trees within a growing stand will greatly influence CMAI.
Conifer trees appropriate for site index measurement typically occur in community phases 1.3. They are selected according to guidance listed in the site index publications.
Table 10. Representative site productivity
| Common name | Symbol | Site index low | Site index high | CMAI low | CMAI high | Age of CMAI | Site index curve code | Site index curve basis | Citation |
|---|---|---|---|---|---|---|---|---|---|
| white fir | ABCO | 50 | 98 | 91 | 234 | 70 | 30 | – | |
| white fir | ABCO | 50 | 98 | 91 | 233 | – | – | 100TA | Meyer, Walter H. 1961. Yield of even-aged stands of ponderosa pine. USDA Technical Bulletin 630. (1938 version revised in 1961). |
| Jeffrey pine | PIJE | 98 | 117 | 99 | 136 | – | – | 100TA | Meyer, Walter H. 1961. Yield of even-aged stands of ponderosa pine. USDA Technical Bulletin 630. (1938 version revised in 1961). |
| Jeffrey pine | PIJE | 98 | 117 | 99 | 135 | 40 | 600 | – |
Supporting information
Inventory data references
The following NRCS plots were used to describe this ecological site:
Community 1.1
789247- type location
Community 1.1 to 1.5 transition
789169
789287
789303
Community 1.2
789310
Community 1.4
789351
Community 1.5
789289
789311
Type locality
| Location 1: Plumas County, CA | |
|---|---|
| Township/Range/Section | T30 N R5 E S23 |
| UTM zone | N |
| UTM northing | 4478291 |
| UTM easting | 635986 |
| General legal description | The site location is approx. 0.4 miles east of Drakesbad Guest Ranch, on the south facing slope. |
Other references
Azuma, David L; Donnegan, Joseph; and Gedney, Donald (2004). Southwest Oregon Biscuit Fire: An alalysis of Forest Resources and Fire Severity. U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station, Research Paper PNW-RP-560
Beaty, Matthew and Taylor, Alan H. (2001). Spatial and Temporal Variation of Fire Regimes in a Mixed Conifer Forest Landscape, Southern Cascades, California, USA. Journal of Biogeography, 28, 955-966.
Bekker, Mathew F. and Taylor, Alan H. (2001). Gradient Analysis of Fire Regimes in Montane Forest of the Southern Cascade Range, Thousand Lakes Wilderness, California, USA. Plant Ecology 155: 15-23.
Bohne, Michael (eds.) (2006). California Forest Pest Conditions – 2006. Forest Health Protection, USDA Forest Service, Pacific Southwest Region in cooperation with other member organizations. California Forest Pest Council.
Gucker, Corey L. (2007). Pinus jeffreyi. In: Fire Effects Information System, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer). Available: http://www.fs.fed.us/database/feis/ [ 2008, March 5].
Habeck, R. J. 1992. Calocedrus decurrens. In: Fire Effects Information System, [Online].
U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer).
Available: http://www.fs.fed.us/database/feis/ [2008, March 27].
Heath, James P. (1967). Primary Conifer Succession, Lassen Volcanic National Park. Ecology, Vol. 48, No. 2. (Mar., 1967), pp. 270-275.
Jenkinson, James L., (1990). Pinus jeffreyi Grev. & Balf. Jeffrey Pine. In. Burns, Russell M; Honkala, Barbara H.; [Technical coordinators] 1990. Silvics of North America: Volume 1. Conifers. United States Department of Agriculture (USDA), Forest Service, Agriculture Handbook 54.
Kroh, Glenn C.; White, Joseph D.; Heath, Shelly K.; Pinder III, John E. (2000). Colonization of a Volcanic Mudflow by an Upper Montane Coniferous Forest at Lassen Volcanic National Park, California. American Midland Naturalist, Vol. 143, No. 1. (Jan., 2000), pp. 126-140.
Laacke, Robert J. (1990). Abies concolor (Gord. & Glend) Lindl. Ex Hildebr. White Fir. In. Burns, Russell M; Honkala, Barbara H.; [Technical coordinators] 1990. Silvics of North America: Volume 1. Conifers. United States Department of Agriculture (USDA), Forest Service, Agriculture Handbook 54.
Meyer, Walter H. 1961. Yield of even-aged stands of ponderosa pine. USDA Technical Bulletin 630. (revised 1961). NASIS ID 600
Powers, Robert F.; Oliver, William W. (1990). Libocedrus decurrens Torr. incense-cedar. In: Burns, Russell M.; Honkala, Barbara H., technical coordinators. Silvics of North America. Volume 1. Conifers. Agric. Handb. 654. Washington, DC: U.S. Department of Agriculture, Forest Service: 173-180. [13382]
Reeves, Sonja L. 2006. Ceanothus cordulatus. In: Fire Effects Information System, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer). Available: http://www.fs.fed.us/database/feis/ [2009, October 10].
Royce, E. B. and Barbour, M.G, 2001. Mediterranean Climate Effects. I. Conifer Water Use Across a Sierra Nevada Ecotone. American Journal of Botany 88(5): 911–918. 2001.
Royce, E. B. and Barbour, M.G, 2001. Mediterranean Climate Effects. II. Conifer Growth Phenology Across a Sierra Nevada Ecotone. American Journal of Botany 88(5): 919–932. 2001.
Schumacher, Francis X. 1926.Yield, stand, and volume tables for white fir in the California pine region. University of California Agricultural Experiment Station Bulletin 407. NASIS ID 030
Skinner, Carl N. and Chang Chi-Ru, (1996). Fire Regimes, Past and Present. Sierra Nevada Ecosystems Project: Final Report to Congress, Vol 2, Assessments and scientific basis for management options. Davis: University of California, Centers for Water and Wildland Resources. Chapter 38, p. 1041.
Smith, Sydney (1994). Ecological Guide to Eastside Pine Associations. USDA Forest Service, Pacific Southwest Region, R5-ECOL-TP-004.
Taylor, A. H. (2000). Fire Regimes and Forest Changes in Mid and Upper Montane Forest of the Southern Cascades, Lassen Volcanic National Park, California, U.S.A. Journal of Biogeography, 27, 87-104.
Taylor, Alan H. and Solem, Michael N. (2001). Fire Regimes and Stand Dynamics in an Upper Montane Forest Landscape in the Southern Cascades, Caribou Wilderness, California. Journal of the Torrey Botanical Society, Vol. 128, No. 4. (Oct. - Dec., 2001), pp. 350-361.
USDA, (2003). Forest Insect Conditions, Forest Pest Conditions in California -2003, 2003. http://www.fs.fed.us/r5/spf/ publications/cond2003/4-2003rpt-insects.pdf
Zouhar, Kris (2001). Abies concolor. In: Fire Effects Information System, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer). Available: http://www.fs.fed.us/database/feis/ [ 2008, March 5].
Contributors
Lyn Townsend
Marchel M. Munnecke
Rangeland health reference sheet
Interpreting Indicators of Rangeland Health is a qualitative assessment protocol used to determine ecosystem condition based on benchmark characteristics described in the Reference Sheet. A suite of 17 (or more) indicators are typically considered in an assessment. The ecological site(s) representative of an assessment location must be known prior to applying the protocol and must be verified based on soils and climate. Current plant community cannot be used to identify the ecological site.
| Author(s)/participant(s) | |
|---|---|
| Contact for lead author | |
| Date | |
| Approved by | |
| Approval date | |
| Composition (Indicators 10 and 12) based on | Annual Production |
Indicators
-
Number and extent of rills:
-
Presence of water flow patterns:
-
Number and height of erosional pedestals or terracettes:
-
Bare ground from Ecological Site Description or other studies (rock, litter, lichen, moss, plant canopy are not bare ground):
-
Number of gullies and erosion associated with gullies:
-
Extent of wind scoured, blowouts and/or depositional areas:
-
Amount of litter movement (describe size and distance expected to travel):
-
Soil surface (top few mm) resistance to erosion (stability values are averages - most sites will show a range of values):
-
Soil surface structure and SOM content (include type of structure and A-horizon color and thickness):
-
Effect of community phase composition (relative proportion of different functional groups) and spatial distribution on infiltration and runoff:
-
Presence and thickness of compaction layer (usually none; describe soil profile features which may be mistaken for compaction on this site):
-
Functional/Structural Groups (list in order of descending dominance by above-ground annual-production or live foliar cover using symbols: >>, >, = to indicate much greater than, greater than, and equal to):
Dominant:
Sub-dominant:
Other:
Additional:
-
Amount of plant mortality and decadence (include which functional groups are expected to show mortality or decadence):
-
Average percent litter cover (%) and depth ( in):
-
Expected annual annual-production (this is TOTAL above-ground annual-production, not just forage annual-production):
-
Potential invasive (including noxious) species (native and non-native). List species which BOTH characterize degraded states and have the potential to become a dominant or co-dominant species on the ecological site if their future establishment and growth is not actively controlled by management interventions. Species that become dominant for only one to several years (e.g., short-term response to drought or wildfire) are not invasive plants. Note that unlike other indicators, we are describing what is NOT expected in the reference state for the ecological site:
-
Perennial plant reproductive capability:
Print Options
Sections
Font
Other
The Ecosystem Dynamics Interpretive Tool is an information system framework developed by the USDA-ARS Jornada Experimental Range, USDA Natural Resources Conservation Service, and New Mexico State University.
Click on box and path labels to scroll to the respective text.