Natural Resources
Conservation Service
Ecological site R025XY042NV
SHALLOW LOAM 14-16 P.Z.
Last updated: 4/25/2024
Accessed: 04/26/2025
General information
Provisional. A provisional ecological site description has undergone quality control and quality assurance review. It contains a working state and transition model and enough information to identify the ecological site.
MLRA notes
Major Land Resource Area (MLRA): 025X–Owyhee High Plateau
MLRA Notes 25—Owyhee High Plateau
This area is in Nevada (56 percent), Idaho (30 percent), Oregon (12 percent), and Utah (2 percent). It makes up about 27,443 square miles. MLRA 25 is characteristically cooler and wetter than the neighboring MLRAs of the Great Basin. The western boundary is marked by a gradual transition to the lower and warmer basins of MLRA 24. The boundary to the south-southeast, with MLRA 28B, is marked by gradual changes in geology marked by an increased dominance of singleleaf pinyon and Utah juniper and a reduced presence of Idaho fescue. The boundary to the north, with MLRA 11, is a rapid transition from the lava plateau topography to the lower elevation Snake River Plain.
Physiography:
All of this area lies within the Intermontane Plateaus. The southern half is in the Great Basin section of the Basin and Range province. This part of the MLRA is characterized by isolated, uplifted fault-block mountain ranges separated by narrow, aggraded desert plains. This geologically older terrain has been dissected by numerous streams draining to the Humboldt River.
The northern half of the area lies within the Columbia Plateaus province. This part of the MLRA forms the southern boundary of the extensive Columbia Plateau basalt flows. Most of the northern half is in the Payette section, but the northeast corner is in the Snake River Plain section. Deep, narrow canyons draining into the Snake River have been incised into this broad basalt plain. Elevation ranges from 3,000 to 7,550 feet on rolling plateaus and in gently sloping basins. It is more than 9,840 feet on some steep mountains. The Humboldt River crosses the southern half of this area
Geology:
The dominant rock types in this MLRA are volcanic. They include andesite, basalt, tuff, and rhyolite. In the north and west parts of the area, Cretaceous granitic rocks are exposed among Miocene volcanic rocks in mountains. A Mesozoic igneous and metamorphic rock complex dominates the south and east parts of the area. Upper and Lower Paleozoic calcareous sediments, including oceanic deposits, are exposed with limited extent in the mountains. Alluvial fan and basin fill sediments occur in the valleys.
Climate:
The average annual precipitation in most of this area is typically 11 to 22 inches. It increases to as much as 49 inches at the higher elevations. Rainfall occurs in spring and sporadically in summer. Precipitation occurs mainly as snow in winter. The precipitation is distributed fairly evenly throughout fall, winter, and spring. The amount of precipitation is lowest from midsummer to early autumn. The average annual temperature is 33 to 51 degrees F. The freeze-free period averages 130 days and ranges from 65 to 190 days, decreasing in length with elevation. It is typically less than 70 days in the mountains.
Water:
The supply of water from precipitation and streamflow is small and unreliable, except along the Owyhee, Bruneau, and Humboldt Rivers. Streamflow depends largely on accumulated snow in the mountains. Surface water from mountain runoff is generally of excellent quality and suitable for all uses. The basin fill sediments in the narrow alluvial valleys between the mountain ranges provide some ground water for irrigation. The alluvial deposits along the large streams have the most ground water. Based on measurements of water quality in similar deposits in adjacent areas, the basin fill deposits probably contain moderately hard water. The water is suitable for almost all uses. The carbonate rocks in this area are considered aquifers, but they are little used. Springs are common along the edges of the limestone outcrops.
Soils:
The dominant soil orders in this MLRA are Aridisols and Mollisols. The soils in the area dominantly have a mesic or frigid temperature regime and an aridic, aridic bordering on xeric, or xeric moisture regime. Soils with aquic moisture regimes are limited to drainage or spring areas, where moisture originates or runs on and through. These soils are of a very limited extent throughout the MLRA. They generally are well drained, clayey or loamy, and shallow or moderately deep. Most of the soils formed in mixed parent material. Volcanic ash and loess mantle the landscape. Surface soil textures are loam and silt loam with ashy texture modifiers in some areas. Argillic horizons occur on the more stable landforms. They are exposed nearer the soil surface on convex landforms, where ash and loess deposits are more likely to erode. Soils that formed in carbonatic parent material in areas that receive less than 12 inches of precipitation are characterized by calcic horizons throughout the profile, while soils in areas that receive more than 12 inches of precipitation do not have calcic horizons in the upper part of the profile. Soils that formed on stable landforms at the lower elevations are dominated by ochric horizons. Soils that formed at the middle and upper elevations are characterized by mollic epipedons. Soils in drainage areas at all elevations that receive moisture running on or through them are characterized by thicker mollic epipedons.
Biological Resources:
This MLRA supports shrub-grass vegetation. Lower elevations are characterized by Wyoming big sagebrush associated with bluebunch wheatgrass, western wheatgrass, and Thurber’s needlegrass. Other important plants include bluegrass, squirreltail, penstemon, phlox, milkvetch, lupine, Indian paintbrush, aster, and rabbitbrush. Black sagebrush occurs but is less extensive. Singleleaf pinyon and Utah juniper occur in limited areas. With increasing elevation and precipitation, vast areas characterized by mountain big sagebrush or low sagebrush/early sagebrush in association with Idaho fescue, bluebunch wheatgrass, needlegrasses, and bluegrass become common. Snowberry, curl-leaf mountain mahogany, ceanothus, and juniper also occur. Mountains at the highest elevations support whitebark pine, Douglas-fir, limber pine, Engelmann spruce, subalpine fir, aspen, and curl-leaf mountain mahogany.
Major wildlife species include mule deer, bighorn sheep, pronghorn, mountain lion, coyote, bobcat, badger, river otter, mink, weasel, golden eagle, red-tailed hawk, ferruginous hawk, Swainson’s hawk, northern harrier, prairie falcon, kestrel, great horned owl, short-eared owl, long-eared owl, burrowing owl, pheasant, sage grouse, chukar, gray partridge, and California quail. Reptiles and amphibians include western racer, gopher snake, western rattlesnake, side-blotched lizard, western toad, and spotted frog. Fish species include bull, red band, and rainbow trout.
Ecological site concept
This site occurs on mountain summits, mountain shoulders and steep sideslopes on all aspects. Slopes range from 15 to 75 percent, but slope gradients of 30 to 75 percent are most typical. Elevations range from 6,500 to 9,000 feet
The soils associated with this site are shallow to bedrock and well drained. Depth to a moderately fine or fine textured subsoil is normally less than 10 inches. The soils have 35 to 75 percent coarse fragments throughout the profile. They have a high amount of gravels, cobbles and stones on the surface which occupy plant growing space yet provide a stabilizing affect on surface erosion conditions.
The reference plant community is dominated by bluebunch wheatgrass, Idaho fescue and mountain big sagebrush.
Associated sites
| R025XY004NV |
LOAMY SLOPE 16+ P.Z. |
|---|---|
| R025XY016NV |
SOUTH SLOPE 14-18 P.Z. |
| R025XY017NV |
CLAYPAN 12-16 P.Z. |
| R025XY071NV |
MAHOGANY SAVANNA 14-16 P.Z. |
Similar sites
| R025XY007NV |
GRAVELLY LOAM 12-16 P.Z. More productive site; PUTR2 dominant shrub |
|---|---|
| R025XY012NV |
LOAMY SLOPE 12-16 P.Z. More productive site |
| R025XY016NV |
SOUTH SLOPE 14-18 P.Z. More productive site; LECI4-BRMA4 codominant grasses |
| R025XY009NV |
SOUTH SLOPE 12-14 P.Z. More productive site |
Table 1. Dominant plant species
| Tree |
Not specified |
|---|---|
| Shrub |
(1) Artemisia tridentata subsp. vaseyana |
| Herbaceous |
(1) Pseudoroegneria spicata |
Physiographic features
This site occurs on mountain summits, mountain shoulders and steep sideslopes on all aspects. Slopes range from 15 to 75 percent, but slope gradients of 30 to 75 percent are most typical. Elevations are 6500 to 9000 feet
Table 2. Representative physiographic features
| Landforms |
(1)
Mountains
> Mountain slope
|
|---|---|
| Runoff class | High to very high |
| Flooding frequency | None |
| Ponding frequency | None |
| Elevation | 1,981 – 2,743 m |
| Slope | 15 – 75% |
| Water table depth | 183 cm |
| Aspect | Aspect is not a significant factor |
Climatic features
The climate associated with this site is semiarid, characterized by cold, moist winters and warm, dry summers.
The average annual precipitation ranges from 14 or more inches. Mean annual air temperature is typically <45 degrees F.
Mean annual precipitation across the range in which this ES occurs is 18.58".
Monthly mean precipitation: January 1.65”; February 1.68”; March 1.98”; April 2.43”; May 2.41”; June 1.62”; July 0.61”; August 0.63”; September 0.84”; October 1.41”; November 1.51”; December 1.79”.
*The above data is averaged from the Jarbridge 4N and Lamoille PH WRCC climate stations.
Table 3. Representative climatic features
| Frost-free period (average) | 84 days |
|---|---|
| Freeze-free period (average) | 114 days |
| Precipitation total (average) | 483 mm |
Figure 1. Monthly precipitation range
Figure 2. Monthly average minimum and maximum temperature
Figure 3. Annual precipitation pattern
Figure 4. Annual average temperature pattern
Climate stations used
-
(1) JARBIDGE 7 N [USC00264039], Jackpot, NV
-
(2) LAMOILLE YOST [USC00264394], Spring Creek, NV
Influencing water features
There are no influencing water features associated with this site.
Soil features
The soils associated with this site are shallow to bedrock and well drained. Depth to a moderately fine or fine textured subsoil is normally less than 10 inches. The soils have 35 to 75 percent coarse fragments throughout the profile. They have a high amount of gravels, cobbles and stones on the surface which occupy plant growing space yet provide a stabilizing affect on surface erosion conditions. The available water capacity of these soils is low to very low and runoff is medium. Potential for sheet and rill erosion is slight to moderate depending on slope. The soil series associated with this site include Amene, Onkeyo, Segura, and Wardbay.
A representative soil series is Onkeyo, classified as a loamy-skeletal, mixed, active, frigid Lithic Calcixeroll. These soils are shallow, well drained soil and formed in residuum and colluvium derived from limestone and dolomite. Reaction is slightly alkaline through strongly alkaline and is violently effervescent. Diagnostic horizons include a mollic epipedon that occurs at the soil surface to 8 inches and a calcic horizon that occurs from 8 inches to 15 inches. Clay content in the particle-size control section is 25 to 35 percent. Rock fragments range from 50 to 80 percent, mainly cobbles. Lithology of fragments is mainly limestone and dolostone.
Table 4. Representative soil features
| Parent material |
(1)
Residuum
–
volcanic rock
|
|---|---|
| Surface texture |
(1) Very gravelly silt loam (2) Gravelly loam (3) Very stony sandy clay loam |
| Family particle size |
(1) Loamy-skeletal |
| Drainage class | Well drained |
| Permeability class | Moderately slow to moderate |
| Depth to restrictive layer | 18 – 152 cm |
| Soil depth | 18 – 152 cm |
| Surface fragment cover <=3" | 32 – 38% |
| Surface fragment cover >3" | 21 – 45% |
| Available water capacity (0-101.6cm) |
3.56 – 7.11 cm |
| Calcium carbonate equivalent (0-101.6cm) |
1 – 60% |
| Electrical conductivity (0-101.6cm) |
0 – 2 mmhos/cm |
| Sodium adsorption ratio (0-101.6cm) |
0 |
| Soil reaction (1:1 water) (0-101.6cm) |
6.6 – 9 |
| Subsurface fragment volume <=3" (Depth not specified) |
0 – 23% |
| Subsurface fragment volume >3" (Depth not specified) |
7 – 36% |
Ecological dynamics
An ecological site is the product of all the environmental factors responsible for its development and has a set of key characteristics that influence a site’s resilience to disturbance and resistance to invasives. Key characteristics include 1) climate (precipitation and temperature), 2) topography (aspect, slope, elevation, and landform), 3) hydrology (infiltration and runoff), 4) soils (depth, texture, structure, and organic matter), 5) plant communities (functional groups and productivity), and 6) natural disturbance regime (fire, herbivory, etc.) (Caudle et al. 2013). Biotic factors that influence resilience include site productivity, species composition and structure, and population regulation and regeneration (Chambers et al. 2013).
This ecological site is dominated by deep-rooted cool season, perennial bunchgrasses and long-lived shrubs (50+ years) with high root to shoot ratios. The dominant shrubs usually root to the full depth of the winter-spring soil moisture recharge, which ranges from 1.0 to over 3.0 meters (Dobrowolski et al. 1990). Root length of mature sagebrush plants was measured to a depth of 2 meters in alluvial soils in Utah (Richards and Caldwell 1987). Tap roots of antelope bitterbrush have been documented from 4.5 to 5.4 meters in length (McConnell 1961). These shrubs have a flexible generalized root system with development of both deep taproots and laterals near the surface (Comstock and Ehleringer 1992).
Periodic drought regularly influences sagebrush ecosystems and drought duration and severity has increased throughout the 20th century in much of the Intermountain West. Major shifts away from historical precipitation patterns have the greatest potential to alter ecosystem function and productivity. Species composition and productivity can be altered by the timing of precipitation and water availability within the soil profile (Bates et al. 2006).
Mountain big sagebrush and antelope bitterbrush are generally long-lived; therefore it is unnecessary for new individuals to recruit every year for perpetuation of the stand. Simultaneous low, continuous recruitment and infrequent large recruitment events are the foundation of population maintenance (Noy-Meir 1973). Survival of the seedlings is dependent upon adequate moisture conditions.
Native insect outbreaks are also important drivers of ecosystem dynamics in sagebrush communities. Climate is generally believed to influence the timing of insect outbreaks especially with regard to Aroga moth (Aroga websteri), a sagebrush defoliator. Aroga moth infestations have occurred in the Great Basin in the 1960s, early 1970s, and have been ongoing in Nevada since 2004 (Bentz et al 2008). Thousands of acres of big sagebrush have been impacted, with partial to complete die-off observed in both individual plants and entire stands (Furniss and Barr 1975).
The Great Basin sagebrush communities have high spatial and temporal variability in precipitation both among years and within growing seasons. Nutrient availability is typically low but increases with elevation and closely follows moisture availability. The invasibility of plant communities is often linked to resource availability. Disturbance can decrease resource uptake due to damage or mortality of the native species and depressed competition. It can also increase resource pools via the decomposition of dead plant material following disturbance. The invasion of sagebrush communities by cheatgrass (Bromus tectorum) has been linked to disturbances (fire, abusive grazing) that have resulted in fluctuations in resources (Chambers et al. 2007).
The perennial bunchgrasses that are co-dominant with the shrubs include bluebunch wheatgrass and Idaho fescue. These species generally have somewhat shallower root systems than the shrubs, but root densities are often as high as or higher than those of shrubs in the upper 0.5 m of the soil profile. The root systems of short lived perennial grasses such as Sandberg bluegrass and mountain brome penetrate only the upper 40cm of the soil, whereas longer lived perennial bunchgrasses can reach depths up to 160 cm (Spence 1937). General differences in root depth distributions between grasses and shrubs results in resource partitioning in these shrub/grass systems.
South-facing slopes will generally express a higher abundance of bluebunch wheatgrass, while north-facing slopes will contain more Idaho fescue. Production will be higher on sites with deeper soils. Overgrazing by livestock and horses will cause a decrease in deep-rooted perennial bunchgrasses, mainly Idaho fescue and bluebunch wheatgrass. As grass cover declines, the potential for invasion by annual non-native species (such as cheatgrass), singleleaf pinyon (Pinus monophylla) and Utah juniper (Juniperus osteosperma) will increase. Continued inappropriate grazing management may result in an increase in Sandberg bluegrass (Poa secunda), balsamroot, lupine, sagebrush, and rabbitbrush (Chrysothamnus viscidiflorus).
As ecological condition declines, mountain big sagebrush and rabbitbrush become dominant with increases of Sandberg’s bluegrass, bottlebrush squirreltail, phlox and mat-forming forbs in the understory. Cheatgrass is likely to invade this site.
This ecological site has low to high resilience to disturbance and resistance to invasion. Increased resilience increases with elevation, aspect, increased precipitation and increased nutrient availability. Six possible stable states have been identified for the Shallow Loam 14-16" ecological site.
Fire Ecology:
Pre-settlement fire return intervals in mountain big sagebrush communities varies from 15 to 25 years (Burkhardt and Tisdale 1969, Houston 1973, Miller 2000). Mountain big sagebrush is killed by fire (Neuenschwander 1980, Blaisdell et al. 1982), and does not resprout (Blaisdell 1953). Post-fire regeneration occurs from seed and will vary depending on site characteristics, seed source, and fire characteristics. Mountain big sagebrush seedlings can grow rapidly and may reach reproductive maturity within 3 to 5 years (Bunting et al. 1987). Mountain big sagebrush may return to pre-burn density and cover within 15-20 years following fire, but establishment after severe fires may proceed more slowly and can take up to 50 years (Bunting et al. 1987, Ziegenhagen 2003, Miller and Heyerdahl 2008, Ziegenhagen and Miller 2009). The introduction of annual weedy species such as cheatgrass may cause an increase in fire frequency and eventually lead to an annual dominated community. Conversely, without fire, sagebrush will increase and the potential for encroachment by pinyon and juniper also increases. Without fire or changes in management, pinyon and juniper will dominate the site and mountain big sagebrush will be severely reduced. The herbaceous understory will also be reduced; Idaho fescue may remain underneath trees on north facing slopes. The potential for soil erosion increases as the woodland matures and the understory plant community cover declines. Catastrophic wildfire in these tree-controlled sites may lead to an annual weed dominated site.
Antelope bitterbrush is moderately fire tolerant (McConnell and Smith 1977). It regenerates by seed and resprouting (Blaisdell and Mueggler 1956, McArthur et al. 1982), however sprouting ability is highly variable and has been attributed to genetics, plant age, phenology, soil moisture and texture, and fire severity (Blaisdell and Mueggler 1956, Blaisdell et al. 1982, Clark et al. 1982, Cook et al. 1994). Bitterbrush sprouts from a region on the stem approximately 1.5 inches above and below the soil surface; the plant rarely sprouts if the root crown is killed by fire (Blaisdell and Mueggler 1956). Low intensity fires may allow for bitterbrush to sprout; however, community response also depends on soil moisture levels at time of fire (Murray 1983). Lower soil moisture allows more charring of the stem below ground level (Blaisdell and Mueggler 1956), thus sprouting will usually be more successful after a spring fire than after a fire in summer or fall (Murray 1983, Busse et al. 2000, Kerns et al. 2006). If cheatgrass is present, bitterbrush seedling success is much lower. The factor that most limits establishment of bitterbrush seedlings is competition for water resources with cheatgrass, an invasive species (Clements and Young 2002).
Depending on fire severity, rabbitbrush, Utah serviceberry (Amelanchier utahensis) and mountain snowberry (Symphoricarpos orbiculatus) may increase after fire. Mountain snowberry is also top-killed by fire, but resprouts after fire from rhizomes (Leege and Hickey 1971, Noste and Bushey 1987). Snowberry has been noted to regenerate well and exceed pre-burn biomass in the third season after a fire (Merrill et al. 1982). Utah serviceberry resprouts from the root crown. Douglas’ rabbitbrush is top-killed by fire, but sprouts vigorously after fire (Kuntz 1982, Akinsoji 1988). As cheatgrass increases, fire frequencies also increase to between 0.23 and 0.43 times a year; at this rate, even sprouting rabbitbrush will not survive (Whisenant 1990).
The effect of fire on bunchgrasses relates to culm density, culm-leaf morphology, and the size of the plant. The initial condition of bunchgrasses within the site, coupled with seasonality and intensity of the fire factor into individual species’ responses. For most forbs and grasses, the growing points are located at or below the soil surface providing relative protection from disturbances which decrease above ground biomass, such as grazing or fire. Thus, fire mortality is more correlated to duration and intensity of heat which is related to culm density, culm-leaf morphology, size of plant and abundance of old growth (Wright 1971, Young 1983).
Mountain brome is a robust, coarse-stemmed, short lived perennial bunchgrass that can grow from 1 to 5 feet in height (Dayton 1937, Tilley et al. 2004). It is commonly seeded after wildfires due to its ability to establish quickly and reduce erosion (Tilley et al. 2004). Mountain brome significantly decreases after burning (Nimir and Payne 1978).
Bluebunch wheatgrass has coarse stems with little leafy material, therefore the aboveground biomass burns rapidly and little heat is transferred downward into the crowns (Young 1983). Bluebunch wheatgrass was described as fairly tolerant of burning, other than in May in eastern Oregon (Britton et al. 1990). Uresk et al. (1976) reported burning increased vegetative and reproductive vigor of bluebunch wheatgrass. Thus, bluebunch wheatgrass is considered to experience slight damage to fire but is more susceptible in drought years (Young 1983). Most authors classify the plant as undamaged by fire (Kuntz 1982).
Basin wildrye is relatively resistant to fire, particularly dormant season fire, as plants sprout from surviving root crowns and rhizomes (Zschaechner 1985). Miller et al. (2013) reported increased total shoot and reproductive shoot densities in the first year following fire; by year two, however, there was little difference between burned and control treatments.
Idaho fescue’s response to fire varies with condition and size of the plant, season and severity of fire, and ecological conditions. Mature Idaho fescue plants are commonly reported to be severely damaged by fire in all seasons (Wright et al. 1979). Initial mortality may be high (in excess of 75%) on severe burns, but usually varies from 20 to 50% (Barrington et al 1988). Rapid burns have been found to leave little damage to root crowns, and new tillers are produced with onset of fall moisture (Johnson et al. 1994). However, Wright and others (1979) found the dense, fine leaves of Idaho fescue provided enough fuel to burn for hours after a fire had passed, thereby killing or seriously injuring the plant regardless of the intensity of the fire (Wright et al. 1979). Idaho fescue is commonly reported to be more sensitive to fire than the other prominent grass on this site, bluebunch wheatgrass (Conrad and Poulton 1966). Robberecht and Defosse (1995) suggested the latter was more sensitive, however. They observed culm and biomass reduction with moderate fire severity in bluebunch wheatgrass, whereas a high fire severity was required for this reduction in Idaho fescue. Given the same fire severity treatment, post-fire culm production was initiated earlier and more rapidly in Idaho fescue (Robberecht and Defosse 1995).
Western needlegrass is moderately damaged by fire. The recovery time is between 3 and 5 years. Little specific information is available on adaptations of Letterman's needlegrass to fire. It is morphologically similar to Columbia needlegrass, which is only slightly to moderately damaged by fire as it has relatively few culms per clumb; this may help to minimize the amount of subsurface heat transfer and subsequent damage. Season of burn affects the plant's ability to survive a fire, and post-fire regeneration occurs through seeding and tillering.
State and transition model
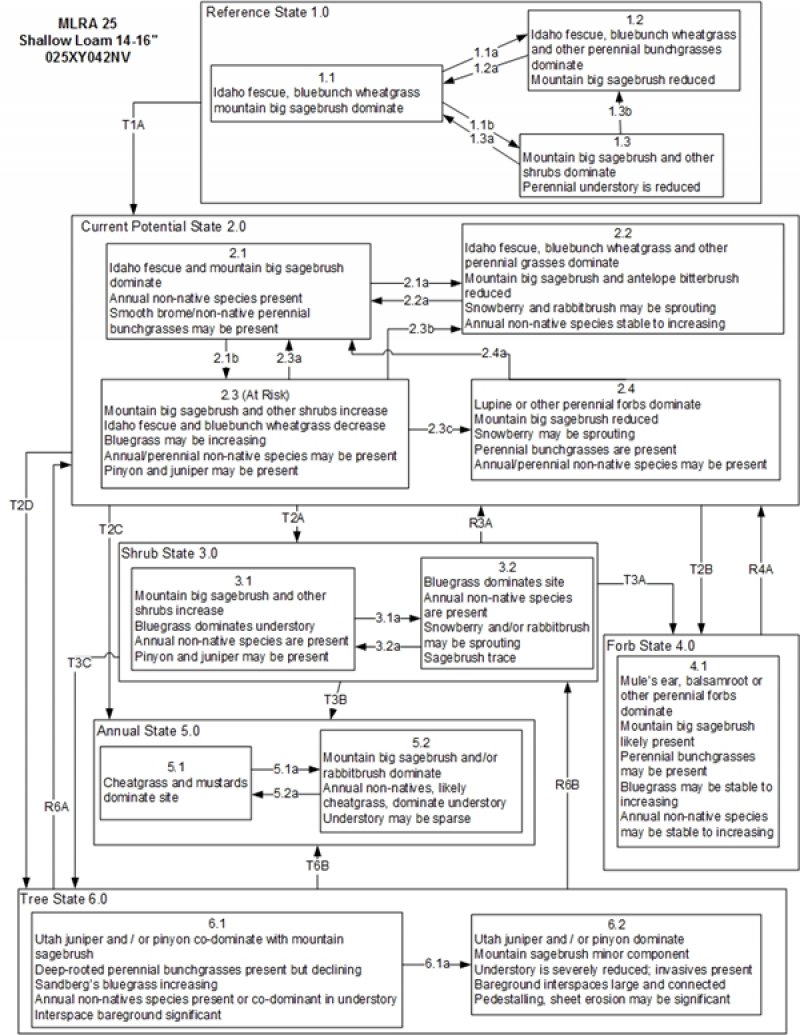
Figure 5. T. Stringham July 2015

Figure 6. T. Stringham July 2015
More interactive model formats are also available.
View Interactive Models
More interactive model formats are also available.
View Interactive Models
Click on state and transition labels to scroll to the respective text
Ecosystem states
States 2, 5 and 6 (additional transitions)
State 1 submodel, plant communities
State 3 submodel, plant communities
State 4 submodel, plant communities
State 5 submodel, plant communities
State 6 submodel, plant communities
State 1
Reference State
The Reference State is a representative of the natural range of variability under pristine conditions. The reference state has three general community phases: a shrub-grass dominant-phase, a perennial grass-dominant phase and a shrub-dominant phase. State dynamics are maintained by interactions between climatic patterns and disturbance regimes. Negative feedbacks enhance ecosystem resilience and contribute to the stability of the state. These include the presence of all structural and functional groups, low fine fuel loads, and retention of organic matter and nutrients. Plant community phase changes are primarily driven by fire, periodic drought and/or insect or disease attack.
Community 1.1
Perennial bunchgrasses/mountain big sagebrush
The reference plant community is dominated by bluebunch wheatgrass, Idaho fescue and mountain big sagebrush. Potential vegetative composition is about 55 percent grasses, 10 percent forbs and 35 percent shrubs. Approximate ground cover (basal and crown) is 10 to 25 percent.
Figure 7. Annual production by plant type (representative values) or group (midpoint values)
Table 5. Annual production by plant type
| Plant type | Low (kg/hectare) |
Representative value (kg/hectare) |
High (kg/hectare) |
|---|---|---|---|
| Grass/Grasslike | 185 | 432 | 616 |
| Shrub/Vine | 118 | 275 | 392 |
| Forb | 34 | 78 | 112 |
| Total | 337 | 785 | 1120 |
Community 1.2
Herbaceous
This community phase is characteristic of a post-disturbance, early seral community phase. Idaho fescue, bluebunch wheatgrass and other perennial grasses dominate. Douglas rabbitbrush, mountain snowberry and Utah serviceberry may be resprouting. Big sagebrush is killed by fire, therefore decreasing within the burned community. Depending on fire severity or intensity of Aroga moth infestations, patches of intact sagebrush may remain. Perennial forbs may increase post-fire but will likely return to pre-burn levels within a few years.
Community 1.3
Mountain big sagebrush/perennial bunchgrasses
Mountain big sagebrush increases in the absence of disturbance. Decadent sagebrush dominates the overstory and the deep-rooted perennial bunchgrasses in the understory are reduced either from competition with shrubs and/or from herbivory. Sandberg bluegrass and/or squirreltail will likely increase in the understory and may be the dominant grass on the site.
Pathway 1.1a
Community 1.1 to 1.2
Fire will decrease or eliminate the overstory of sagebrush and allow for the perennial bunchgrasses to dominate the site. Fires will typically be low severity resulting in a mosaic pattern due to low fuel loads. A fire following an unusually wet spring or a change in management may be more severe and reduce sagebrush cover to trace amounts. A severe infestation of Aroga moth could also cause a large decrease in sagebrush within the community, giving a competitive advantage to the perennial grasses and forbs.
Pathway 1.1b
Community 1.1 to 1.3
Time and lack of disturbance such as fire allows for sagebrush to increase and become decadent. Long-term drought, herbivory, or combinations of these will cause a decline in perennial bunchgrasses and fine fuels leading to a reduced fire frequency allowing big sagebrush to dominate the site.
Pathway 1.2a
Community 1.2 to 1.1
Absence of disturbance over time and appropriate herbivory management
Pathway 1.3a
Community 1.3 to 1.1
A low severity fire, Aroga moth or combinations will reduce the sagebrush overstory and create a sagebrush/grass mosaic with sagebrush and perennial bunchgrasses codominant.
Pathway 1.3b
Community 1.3 to 1.2
Fire will decrease or eliminate the overstory of sagebrush and allow for the perennial bunchgrasses to dominate the site. Fires will typically be low severity due to low fine fuel loads. A fire following an unusually wet spring or a change in management may be more severe and reduce sagebrush cover to trace amounts. A severe infestation of Aroga moth could also cause a large decrease in sagebrush within the community, giving a competitive advantage to the perennial grasses and forbs.
State 2
Current Potential State
This state is similar to the Reference State 1.0 with four similar community phases. Ecological function has not changed, however the resiliency of the state has been reduced by the presence of invasive weeds. Non-natives may increase in abundance but will not become dominant within this State. These non-natives can be highly flammable, and can promote fire where historically fire had been infrequent. Negative feedbacks enhance ecosystem resilience and contribute to the stability of the state. These include the presence of all structural and functional groups, low fine fuel loads and retention of organic matter and nutrients. Positive feedbacks decrease ecosystem resilience and stability of the state. These include the non-natives’ high seed output, persistent seed bank, rapid growth rate, ability to cross pollinate and adaptations for seed dispersal.
Community 2.1
Perennial bunchgrasses/mountain big sagebrush/annual non-native species present
This community phase is similar to the Reference State Community Phase 1.1, with the presence of non-native species in trace amounts. The plant community is dominated by Idaho fescue, bluebunch wheatgrass and basin wildrye. Mountain big sagebrush is the dominant shrub with antelope bitterbrush also common on this site. Smooth brome or other perennial non-native bunchgrasses may be present. Cheatgrass is the most likely species to invade.
Community 2.2
Herbaceous
This community phase is characteristic of a post-disturbance, early seral community phase where non-native species are present. Idaho fescue, bluebunch wheatgrass and other perennial grasses dominate. Douglas rabbitbrush, mountain snowberry and Utah serviceberry may be resprouting. Depending on fire severity or intensity of Aroga moth infestations, patches of intact sagebrush may remain. Perennial forbs may increase post-fire but will likely return to pre-burn levels within a few years. Annual non-native species are stable or increasing within the community.
Community 2.3
Mountain big sagebrush/perennial bunchgrasses (at risk)
Mountain big sagebrush, rabbitbrush and bitterbrush increase, Idaho fescue and bluebunch wheatgrass decrease. Sandberg bluegrass may be increasing. Smooth brome and other non-native species are stable to increasing. Juniper and pinyon may be present as a result of encroachment from neighboring sites, and lack of disturbance.
Community 2.4
Forbs
Lupine and other perennial forbs dominate the site. Mountain big sagebrush may be present in trace amounts. Snowberry may be sprouting. Perennial bunchgrasses are present. Smooth brome and/or other perennial non-native bunchgrasses may be present. This community phase can be maintained for several years with spring grazing and poor distribution of grazing animals. With a change in grazing management mountain big sagebrush will increase and perennial forb community will eventually decrease.
Pathway 2.1a
Community 2.1 to 2.2
Fire will decrease or eliminate the overstory of sagebrush and allow for the perennial bunchgrasses to dominate the site. Fires will typically be small and patchy due to low fuel loads. A fire following an unusually wet spring or a change in management may be more severe and reduce sagebrush cover to trace amounts. A severe infestation of Aroga moth could also cause a large decrease in sagebrush within the community, giving a competitive advantage to the perennial grasses and forbs.
Pathway 2.1b
Community 2.1 to 2.3
Time and lack of disturbance allows for sagebrush to increase and become decadent. Chronic drought will reduce fine fuels and lead to a reduced fire frequency allowing big sagebrush to dominate the site. Inappropriate grazing management will reduce the perennial bunchgrass understory; conversely Sandberg bluegrass may increase in the understory depending on grazing management. Excessive sheep grazing favors Sandberg bluegrass; however, where cattle are the dominant grazers, cheatgrass often increases.
Pathway 2.2a
Community 2.2 to 2.1
Time and lack of disturbance and/or grazing management that favors the establishment and growth of sagebrush allows the shrub component to recover. The establishment of big sagebrush may take many years.
Pathway 2.2b
Community 2.2 to 2.3
Fire
Pathway 2.3a
Community 2.3 to 2.2
A low severity fire, Aroga moth or combinations will reduce the sagebrush overstory and create a sagebrush/grass mosaic.
Pathway a
Community 2.3 to 2.2
A low severity fire, Aroga moth or combinations will reduce the sagebrush overstory and create a sagebrush/grass mosaic.
Pathway 2.3c
Community 2.3 to 2.4
High severity fire following inappropriate grazing management (spring grazing suppresses perennial bunchgrass response) increases the forb community response.
Pathway 2.4a
Community 2.4 to 2.1
Time and lack of disturbance combined with grazing management to facilitate perennial bunchgrass production such as fall season grazing.
State 3
Shrub State
This state has two community phases: a mountain big sagebrush-dominated phase and a rabbitbrush-dominated phase. This state is a product of many years of heavy grazing during time periods harmful to perennial bunchgrasses. Sandberg bluegrass will increase with a reduction in deep rooted perennial bunchgrass competition and become the dominant grass on this site. Sagebrush dominates the overstory and rabbitbrush may be a significant component. Sagebrush cover exceeds site concept and may be decadent, reflecting stand maturity and lack of seedling establishment due to competition with mature plants.
Community 3.1
Mountain big sagebrush/bluegrass
This site has crossed a biotic threshold and site processes (soil hydrology, nutrient cycling, and energy capture) are being controlled by the shrub component of the plant community along with Sandberg bluegrass in the understory. Decadent big sagebrush and rabbitbrush dominate overstory. Deep-rooted perennial bunchgrasses may be present in trace amounts or absent from the community. Sandberg bluegrass and annual non-native species increase, and the amount of bare ground increases. Utah juniper may be present as a result of encroachment from neighboring sites and lack of natural fire. Singleleaf pinyon may be invading.
Community 3.2
Bluegrass/annual non-native species present
Bluegrass dominates the site; annual non-native species may be present but are not dominant. Trace amounts of sagebrush, snowberry and/or rabbitbrush may be present.
Pathway 3.1a
Community 3.1 to 3.2
Fire, heavy fall grazing causing mechanical damage to shrubs, and/or brush treatments with minimal soil disturbance, will greatly reduce the overstory shrubs to trace amounts and allow for Sandberg bluegrass to dominate the site.
Pathway 3.2a
Community 3.2 to 3.1
Absence of disturbance over time will allow for the sagebrush and other shrubs to recover.
State 4
Forb State
This state has one community phase that is dominated by deep-rooted perennial forbs. This state may be the result of many years of abusive grazing allowing the perennial forbs to increase. It may also be compounded by fire. This may occur as “pockets” or inclusions within other states of the same site, and can appear to be localized. The positive feedbacks in this state include the presence of a competitive functional group that possesses deep-rooted taproots and strong lateral roots, the sprouting ability of roots or root crown, high seed production and the ability to monopolize soil moisture.
Community 4.1
Forbs
Mule’s ear (Wyethia amplexicaulis), and/or other perennial forbs dominate the site. Mountain big sagebrush is likely present. Sandberg bluegrass may be stable to increasing, and perennial bunchgrasses are a minor component.
State 5
Annual State
This state has two community phases and is characterized by the dominance of annual non-native species such as cheatgrass and tumble mustard in the understory. Sagebrush and/or rabbitbrush dominate the overstory.
Community 5.1
Annual non-native species
Annual non-native plants such as tumble mustard and cheatgrass dominate the site.
Community 5.2
Shrubs/annual non-native species
Mountain big sagebrush and/or rabbitbrush dominate the overstory. Annual non-native species, likely cheatgrass, dominate the understory. Understory may be sparse.
Pathway 5.1a
Community 5.1 to 5.2
Time and a lack of disturbance will allow for recovery of mountain big sagebrush and/or rabbitbrush. This community phase pathway is unlikely to occur.
Pathway 5.2a
Community 5.2 to 5.1
Fire, brush management, or Aroga moth infestation will reduce or eliminate the sagebrush component on this site and allow for annual non-natives to dominate.
State 6
Tree State
This state has two community phases that are characterized by the dominance of Utah juniper in the overstory. Singleleaf pinyon may also be present. Big sagebrush and perennial bunchgrasses may still be present, but they are no longer controlling site dynamics in this state. Soil moisture, soil nutrients and soil organic matter distribution and cycling have been spatially and temporally altered.
Community 6.1
Trees/annual non-native species
Utah juniper and singleleaf pinyon dominate the overstory. Mountain big sagebrush and rabbitbrush are minor components of the understory. Sandberg bluegrass is present. Deep rooted perennial bunchgrasses are present but declining. Annual non-native species are present or co-dominant in the understory. Pedestalling, sheet, and rill erosion are increasing.
Community 6.2
Trees
Utah juniper and pinyon pine dominate the overstory. The understory is sparse and bare ground may be significant. This community is not likely to change without human interference or a severe wildfire that will transition it to an Annual State 5.0.
Pathway 6.1a
Community 6.1 to 6.2
Utah juniper and pinyon pine dominate the overstory. The understory is sparse and bare ground may be significant. This community is not likely to change without human interference or a severe wildfire that will transition it to an Annual State 5.0.
Transition T1A
State 1 to 2
Trigger: Introduction of annual non-native species Slow variable: Over time the annual non-native plants will increase within the community decreasing organic matter inputs from deep-rooted perennial bunchgrasses resulting in reductions in soil water availability for perennial bunchgrasses. Threshold: Any amount of introduced non-native species causes an immediate decrease in the resilience of the site. Annual non-native species cannot be easily removed from the system and have the potential to significantly alter disturbance regimes from their historic range of variation.
Transition T2A
State 2 to 3
Trigger: To Community Phase 3.1: Inappropriate grazing will decrease or eliminate deep rooted perennial bunchgrasses, increase Sandberg bluegrass and favor shrub growth and establishment. To Community Phase 3.2: Severe fire in community phase 2.3 will remove sagebrush overstory, decrease perennial bunchgrasses and enhance Sandberg bluegrass. Annual non-native species will increase. Slow variables: Long term decrease in deep-rooted perennial grass density resulting in decreased organic matter inputs and reduced soil water. Threshold: Loss of deep-rooted perennial bunchgrasses changes nutrient cycling, nutrient redistribution, and reduces soil organic matter.
Transition T2B
State 2 to 4
Trigger: Inappropriate grazing management and/or fire promote mule ears and other perennial forbs to dominate the site. Persistent spring grazing after a fire will suppress perennial grasses and promote forb production. Slow variable: Increasing density of perennial forbs and soil erosion. Threshold: Loss of deep-rooted perennial bunchgrasses and shrubs changes nutrient capture and cycling within the community and reduced organic matter inputs and soil water.
Transition T2C
State 2 to 5
Trigger: Multiple fires and/or soil disturbing treatments such as drill seeding, roller chopper or Lawson aerator will cause the site to transition to community phase 5.1. Inappropriate grazing management likely by cattle and/or horses can eliminate the native understory and will leave site open for non-native invasive species, which will transition to community phase 5.2. Slow variable: Increased production and cover of non-native annual species. Threshold: Loss of deep-rooted perennial bunchgrasses and shrubs changes nutrient capture and cycling within the community. Increased, continuous fine fuels modify the fire regime by changing intensity, size and spatial variability of fires.
Transition T2D
State 2 to 6
Trigger: Absence of disturbance over time and lack of management action allows for trees to increase and transitions to community phase 6.1. Slow variables: Increased establishment and cover of juniper trees. Threshold: Trees overtop big sagebrush and out-compete shrubs for water and sunlight. Shrub skeletons exceed live shrubs with minimal recruitment of new cohorts.
Restoration pathway R3A
State 3 to 2
Brush management with minimal soil disturbance/seeding of desired species.
Conservation practices
| Brush Management | |
|---|---|
| Range Planting |
Transition T3A
State 3 to 4
Trigger: Fire in combination with inappropriate grazing management can eliminate Sandberg bluegrass understory and transition to 4.1. Slow variable: Increasing density of perennial forbs and soil erosion. Threshold: Loss of deep-rooted perennial bunchgrasses and shrubs changes nutrient capture and cycling within the community and reduced organic matter inputs and reduced soil water.
Transition T3B
State 3 to 5
Trigger: Severe fire will transition to 5.1. Inappropriate grazing management in the presence of annual non-native species will transition to 5.2 Slow variables: Increased production and cover of non-native annual species. Threshold: Increased, continuous fine fuels modify the fire regime by changing intensity, size and spatial variability of fires. Changes in plant community composition and spatial variability of vegetation due to the loss of perennial bunchgrasses and sagebrush truncate energy capture and impact the nutrient cycling and distribution.
Transition T3C
State 3 to 6
Trigger: Time and a lack of disturbance or management action allows for Utah juniper to dominate the site. This may be coupled with grazing management that favors tree establishment by reducing understory herbaceous competition for site resources. Slow variable: Over time the abundance and size of trees will increase resulting in reduced infiltration and increased runoff. Threshold: Trees dominate ecological processes and number of shrub skeletons exceed number of live shrubs.
Restoration pathway R4A
State 4 to 2
Herbicide treatment to reduce perennial forbs may be coupled with seeding of perennial bunchgrasses and other desirable species (Mueggler and Blaisdell 1951).
Conservation practices
| Range Planting | |
|---|---|
| Herbaceous Weed Control |
Restoration pathway R6A
State 6 to 2
This site could be restored with tree removal and seeding or recovery of herbaceous understory. Restoration efforts may result in smooth brome or intermediate wheatgrass after fire. This is most likely to succeed in community phase 6.1.
Conservation practices
| Brush Management | |
|---|---|
| Range Planting |
Restoration pathway R6B
State 6 to 3
Tree removal or fire when Sandberg bluegrass is dominant in the understory will transition to community phase 3.2.
Conservation practices
| Brush Management |
|---|
Transition T6A
State 6 to 5
Trigger: Catastrophic crown fire will reduce or eliminate trees to transition the site to 5.1. Inappropriate tree removal practices such as chop and burn when annual non-natives such as cheatgrass are present will also transition the site to state 5.0. Slow variable: Increased seed production and cover of annual non-native species. Decreased infiltration and increased runoff result in reduction in soil water. Threshold: Increased, continuous fine fuels modify the fire regime by changing intensity, size and spatial variability of fires. Changes in plant community composition and spatial variability of vegetation due to the loss of perennial bunchgrasses and sagebrush truncate energy capture and impact the nutrient cycling and distribution.
Additional community tables
Table 6. Community 1.1 plant community composition
| Group | Common name | Symbol | Scientific name | Annual production (kg/hectare) | Foliar cover (%) | |
|---|---|---|---|---|---|---|
|
Grass/Grasslike
|
||||||
| 1 | Primary Perennial Grasses | 290–612 | ||||
| bluebunch wheatgrass | PSSPS | Pseudoroegneria spicata ssp. spicata | 118–235 | – | ||
| Idaho fescue | FEID | Festuca idahoensis | 118–235 | – | ||
| mountain brome | BRMA4 | Bromus marginatus | 39–78 | – | ||
| basin wildrye | LECI4 | Leymus cinereus | 16–63 | – | ||
| 2 | Secondary Perennial Grasses | 39–78 | ||||
| needlegrass | ACHNA | Achnatherum | 4–24 | – | ||
| bluegrass | POA | Poa | 4–24 | – | ||
|
Forb
|
||||||
| 3 | Perennial Forbs | 39–118 | ||||
| aster | ASTER | Aster | 3–24 | – | ||
| milkvetch | ASTRA | Astragalus | 3–24 | – | ||
| arrowleaf balsamroot | BASA3 | Balsamorhiza sagittata | 3–24 | – | ||
| tapertip hawksbeard | CRAC2 | Crepis acuminata | 3–24 | – | ||
| buckwheat | ERIOG | Eriogonum | 3–24 | – | ||
| lupine | LUPIN | Lupinus | 3–24 | – | ||
| phlox | PHLOX | Phlox | 3–24 | – | ||
|
Shrub/Vine
|
||||||
| 4 | Primary Shrubs | 126–337 | ||||
| mountain big sagebrush | ARTRV | Artemisia tridentata ssp. vaseyana | 78–157 | – | ||
| antelope bitterbrush | PUTR2 | Purshia tridentata | 16–78 | – | ||
| Utah serviceberry | AMUT | Amelanchier utahensis | 16–63 | – | ||
| snowberry | SYMPH | Symphoricarpos | 16–39 | – | ||
| 5 | Secondary Shrubs | 16–63 | ||||
| yellow rabbitbrush | CHVI8 | Chrysothamnus viscidiflorus | 4–16 | – | ||
Interpretations
Animal community
Livestock Interpretations:
This site is suited for livestock grazing. Considerations for grazing management should include timing, intensity and duration of grazing.
Overgrazing leads to an increase in sagebrush and a decline in understory plants like bluebunch wheatgrass . Squirreltail or Sandberg bluegrass will increase temporarily with further degradation. Invasion of annual weedy forbs and cheatgrass could occur with further grazing degradation, leading to a decline in squirreltail and bluegrass and an increase in bare ground. A combination of overgrazing and prolonged drought leads to soil erosion, increased bare ground, and a loss in plant production. Wildlife in sites with cheatgrass present could transition to cheatgrass-dominated communities, and without management, cheatgrass and annual forbs are likely to dominate.
Reduced bunchgrass vigor or density provides an opportunity for Sandberg bluegrass expansion and/or cheatgrass and other invasive species to occupy interspaces, leading to increased fire frequency and potentially an annual plant community. Sandberg bluegrass increases under grazing pressure (Tisdale and Hironaka 1981) and is capable of co-existing with cheatgrass. Excessive sheep grazing favors Sandberg bluegrass; however, where cattle are the dominant grazers, cheatgrass often dominates (Daubenmire 1970). Thus, depending on the season of use, the grazer and site conditions, either Sandberg bluegrass or cheatgrass may become the dominant understory with inappropriate grazing management.
Long-term disturbance response may be influenced by small differences in landscape topography. Concave areas hold a little more moisture and may retain deep-rooted perennial grasses whereas convex areas are slightly less resilient and may have more Sandberg bluegrass present.
Idaho fescue provides important forage for many types of domestic livestock. The foliage cures well and is preferred by livestock in the late fall and winter. Idaho fescue tolerates light to moderate grazing (Ganskopp and Bedell 1980) and is moderately resistant to trampling (Cole 1987). Heavy grazing may lead to replacement of Idaho fescue with non-native species such as cheatgrass (Mueggler 1984).
Mountain brome is ranked as excellent forage for both cattle and horses and good for domestic sheep, though domestic animals will graze mountain brome only when it is fairly succulent. Mountain brome increases with grazing (Leege et al. 1981). A study by Mueggler (1967) found that with clipping, mountain brome increased in herbage production when clipped in June. When clipped in July, mountain brome increased due to reduced competition from forb species. The study also found that after three successive years of clipping, however, mountain brome started to exhibit adverse effects.
Bluebunch wheatgrass is moderately grazing tolerant and is very sensitive to defoliation during the active growth period (Blaisdell and Pechanec 1949, Laycock 1967, Anderson and Scherzinger 1975, Britton et al. 1990). Herbage and flower stalk production was reduced with clipping at all times during the growing season; however, clipping was most harmful during the boot stage (Blaisdell and Pechanec 1949). Tiller production and growth of bluebunch was greatly reduced when clipping was coupled with drought (Busso and Richards 1995). Mueggler (1975) estimated that low vigor bluebunch wheatgrass may need up to 8 years rest to recover. Although an important forage species, it is not always the preferred species by livestock and wildlife.
The early growth and abundant production of basin wildrye make it a valuable source of forage for livestock. It is important for cattle and is readily grazed by both cattle and horses in the early spring and fall. Though coarse-textured during the winter, it may be utilized more frequently by livestock and wildlife when snow has covered low shrubs and other grasses.
Mountain big sagebrush is eaten by domestic livestock but has long been considered to be of low palatability, and a competitor to more desirable species.
Antelope bitterbrush is an important shrub species to domestic livestock (Wood 1995). Antelope bitterbrush is most commonly found on soils which provide minimal restriction to deep root penetration such as coarse textured soil, or finer textured soil with high stone content (Driscoll 1964, Clements and Young 2002). Grazing tolerance of antelope bitterbrush is dependent on site conditions (Garrison 1953).
Stocking rates vary over time depending upon season of use, climate variations, site, and previous and current management goals. A safe starting stocking rate is an estimated stocking rate that is fine-tuned by the client by adaptive management through the year and from year to year.
Wildlife Interpretations:
Basin wildrye provides winter forage for mule deer, though use is often low compared to other native grasses. It also provides summer forage for black-tailed jackrabbits, and because it remains green throughout early summer, it remains available for other small mammal forage for longer time than other grasses. It is intolerant of heavy, repeated, or spring grazing (Krall et al. 1971).
Idaho fescue is an important source of forage for pronghorn and deer in ranges of northern Nevada.
Mountain brome seedheads and seeds provide food for many birds and small mammals. Pronghorn antelope will consume mountain brome primarily in the spring. The palatability of mountain brome is excellent for deer, particularly during the late spring and early summer.
Mountain big sagebrush is a highly preferred winter forage for mule deer: In a study by Personius et al. (1987), mountain big sagebrush was the most preferred sagebrush species. Fecal samples from ungulates in Montana showed that bighorn sheep, mule deer, and elk all consumed mountain big sagebrush in small amounts in winter, while cattle showed no sign of sagebrush use. Reliance on the big sagebrush ecosystem by many wild animals for both food and cover has been documented and reviewed extensively. Sagebrush-grassland communities provide critical sage-grouse breeding and nesting habitats. Meadows surrounded by sagebrush may be used as feeding and strutting grounds. Sagebrush is a crucial component of their diet year-round, and sage-grouse select sagebrush almost exclusively for cover. Sage-grouse prefer mountain big sagebrush and Wyoming big sagebrush communities to basin big sagebrush communities.
Bitterbrush is critical browse for mule deer, as well as domestic livestock, antelope, and elk (Wood 1995).
Hydrological functions
Runoff is medium. Permeability is moderately slow to moderate. Rills are typically none. There may be a few, widely spaced and shallow rills on steeper slopes (30% gradient). Water flow patterns are typically non-existent but can rarely occur on steeper slopes (30% gradient) in areas recently subjected to intense summer convection storms or rapid snowmelt. Pedestals are none to rare. Gullies are non-existent. Perennial herbaceous plants (especially deep-rooted bunchgrasses [i.e. Idaho fescue] slow runoff and increase infiltration. Shrub canopy and associated litter break raindrop impact and provide opportunity for snow catch and accumulation on site.
Recreational uses
Aesthetic value is derived from the diverse floral and faunal composition and the colorful flowering of wild flowers and shrubs during the spring and early summer. This site offers rewarding opportunities to photographers and for nature study. This site is used for camping and hiking and has potential for upland and big game hunting.
Other products
Native peoples used big sagebrush leaves and branches for medicinal teas, and the leaves as a fumigant. Bark was woven into mats, bags and clothing.
Other information
Antelope bitterbrush has been used extensively in land reclamation. Antelope bitterbrush enhances succession by retaining soil and depositing organic material and in some habitats and with some ecotypes, by fixing nitrogen.
Supporting information
Inventory data references
NRCS-RANGE-417 - 2 records
NV-ECS-1 - 1 record
Soils and Physiographic features were gathered from NASIS.
Type locality
| Location 1: Elko County, NV | |
|---|---|
| Latitude | 41° 47′ 18″ |
| Longitude | 115° 40′ 46″ |
| General legal description | NASIS User Site ID: 1994NV007103 |
Other references
Akinsoji, A. 1988. Postfire vegetation dynamics in a sagebrush steppe in southeastern Idaho, USA. Vegetatio 78:151-155.
Anderson, E. W. and R. J. Scherzinger. 1975. Improving quality of winter forage for elk by cattle grazing. Journal of Range Management 28:120-125.
Barrington, M., S. Bunting, and G. Wright. 1988. A Fire Management Plan for Craters of the Moon National Monument. Cooperative Agreement CA-9000-8-0005. Moscow, ID: University of Idaho, Range Resources Department. 52 p. Draft.
Bates, J. D., T. Svejcar, R. F. Miller, and R. A. Angell. 2006. The effects of precipitation timing on sagebrush steppe vegetation. Journal of Arid Environments 64: 670-697.
Bentz, B., D. Alston, and T. Evans. 2008. Great Basin insect outbreaks. In: J. Chambers, N. Devoe, A. Evenden [eds]. Collaborative management and research in the Great Basin -- Examining the issues and developing a framework for action. Gen. Tech. Rep. RMRS-GTR-204. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fort Collins, CO. p. 45-48.
Blaisdell, J. P. 1953. Ecological effects of planned burning of sagebrush-grass range on the Upper Snake River Plains. US Dept. of Agriculture.
Blaisdell, J. P. and W. F. Mueggler. 1956. Sprouting of bitterbrush (Purshia tridentata) following burning or top removal. Ecology 37: 365-370.
Blaisdell, J. P., R. B. Murray, and E. D. McArthur. 1982. Managing intermountain rangelands - sagebrush-grass ranges. USDA Forest Serv. Intermountain Forest and Range Exp. Sta. Gen. Tech. Rep. INT-134.
Blaisdell, J. P. and J. F. Pechanec. 1949. Effects of herbage removal at various dates on vigor of bluebunch wheatgrass and arrowleaf balsamroot. Ecology 30: 298-305.
Bradley, A. F. 1984. Rhizome morphology, soil distribution, and the potential fire survival of eight woody understory species in western Montana. University of Montana.
Britton, C. M., G. R. McPherson, and F. A. Sneva. 1990. Effects of burning and clipping on five bunchgrasses in eastern Oregon. Great Basin Naturalist 50:1 15-120.
Bunting, S. C., B. M. Kilgore, and C. L. Bushey. 1987. Guidelines for prescribed burning sagebrush-grass rangelands in the northern Great Basin. US Department of Agriculture, Forest Service, Intermountain Research Station Ogden, UT, USA.
Burkhardt, J. W. and E. Tisdale. 1969. Nature and successional status of western Juniper vegetation in Idaho. Journal of Range Management:264-270.
Busse, D., A. Simon, and M. Riegel. 2000. Tree-growth and understory responses to low-severity prescribed burning in thinned Pinus ponderosa forests of central Oregon. Forest Science 46: 258-268.
Caudle, D., J. DiBenedetto, M. Karl, H. Sanchez, and C. Talbot. 2013. Interagency Ecological Site Handbook for Rangelands. Available at: http://jornada.nmsu.edu/sites/jornada.nmsu.edu/files/InteragencyEcolSiteHandbook.pdf. Accessed 4 October 2013.
Chambers, J., B. Bradley, C. Brown, C. D’Antonio, M. Germino, J. Grace, S. Hardegree, R. Miller, and D. Pyke. 2013. Resilience to stress and disturbance, and resistance to Bromus tectorum L. invasion in cold desert shrublands of western North America. Ecosystems 17: 1-16.
Chambers, J. C., B. A. Roundy, R. R. Blank, S. E. Meyer, and A. Whittaker. 2007. What makes Great Basin sagebrush ecosystems invasible by Bromus tectorum? Ecological Monographs 77:117-145.
Clark, R. G., M. B. Carlton, and F. A. Sneva. 1982. Mortality of bitterbrush after burning and clipping in eastern Oregon. Journal of Range Management 35: 711-714.
Clements, C. D. and J. A. Young. 2002. Restoring antelope bitterbrush. Rangelands 24: 3-6.
Comstock, J. P. and J. R. Ehleringer. 1992. Plant adaptation in the Great Basin and Colorado plateau. Western North American Naturalist 52:195-215.
Conrad, C. E. and C. E. Poulton. 1966. Effect of a wildfire on Idaho fescue and bluebunch wheatgrass. Journal of Range Management 19:138-141.
Cook, J. G., T. J. Hershey, and L. L. Irwin. 1994. Vegetative response to burning on Wyoming mountain-shrub big game ranges. Journal of Range Management 47: 296-302.
Daubenmire, R. 1970. Steppe vegetation of Washington. 131 pp.
Daubenmire, R. 1975. Plant succession on abandoned fields, and fire influences in a steppe area in southeastern Washington. Northwest Science 49: 36-48.
Driscoll, R. S. 1964. A relict area in the central Oregon juniper zone. Ecology 45:345-353.
Dobrowolski, J. P., M. M. Caldwell, and J. H. Richards. 1990. Basin hydrology and plant root systems. In: C. B. Osmand, L. F. Pitelka, G. M. Hildy [eds]. Plant biology of the basin and range. Ecological Studies. 80: 243-292.
Eckert Jr, R. E. and J. S. Spencer. 1986. Vegetation response of allotments grazed under rest-rotation management. Journal of Range Management: 166-174.
Eckert, R. E., Jr. and J. S. Spencer. 1987. Growth and reproduction of grasses heavily grazed under rest-rotation management. Journal of Range Management 40:156-159.
Fire Effects Information System [Online]. http://www.fs.fed.us/database/feis.
Furniss, M. M. and W. F. Barr. 1975. Insects affecting important native shrubs of the northwestern United States General Technical Report INT-19. Intermountain Forest and Range Experiment Station, U.S. Department of Agriculture, Forest Service. Ogden, UT. p. 68.
Gaffney, W. S. 1941. The effects of winter elk browsing, South Fork of the Flathead River, Montana. The Journal of Wildlife Management 5:427-453.
Ganskopp, D., L. Aguilera, and M. Vavra. 2007. Livestock Forage Conditioning Among Six Northern Great Basin Grasses. Rangeland Ecology & Management 60: 71-78.
Garrison, G. A. 1953. Effects of clipping on some range shrubs. Journal of Range Management 6:309-317.
Houghton, J.G., C.M. Sakamoto, and R.O. Gifford. 1975. Nevada’s weather and climate, special publication 2. Nevada Bureau of Mines and Geology, Mackay School of Mines, University of Nevada, Reno, NV.
Houston, D. B. 1973. Wildfires in northern Yellowstone National Park. Ecology 54:1111-1117.
Johnson, C. G., R. R. Clausnitzer, P. J. Mehringer, and C. Oilver. 1994. Biotic and abiotic processes of eastside ecosystems: The effects of management on plant and community ecology, and on stand and landscape vegetation dynamics. Forest Service General Technical Report. Forest Service, Portland, OR (United States). Pacific Northwest Research Station.
Kasworm, W. F., L. R. Irby, and H. B. I. Pac. 1984. Diets of ungulates using winter ranges in northcentral Montana. Journal of Range Management 37:67-71.
Kerns, B. K., W. G. Thies, and C. G. Niwa. 2006. Season and severity of prescribed burn in ponderosa pine forests: Implications for understory native and exotic plants. Ecoscience 13:44-55.
Krall, J. L., J. R. Stroh, C. S. Cooper, and S. R. Chapman. 1971. Effect of time and extent of harvesting basin wildrye. Journal of Range Management 24: 414-418.
Kuntz, D. E. 1982. Plant response following spring burning Plant in an Artemisia tridentata subsp. vaseyana/Festuca Idahoensis habitat type. University of Idaho.
Laycock, W. A. 1967. How heavy grazing and protection affect sagebrush-grass ranges. Journal of Range Management: 206-213.
Leege, T. A. and W. O. Hickey. 1971. Sprouting of northern Idaho shrubs after prescribed burning. The Journal of Wildlife Management: 508-515.
Majerus, M. E. 1992. High-stature grasses for winter grazing. Journal of Soil and Water Conservation 47: 224-225.
McArthur, E. D., A. Blaner, A. P. Plummer, and R. Stevens. 1982. Characteristics and hybridization of important intermountain shrubs: 3. sunflower family. U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. Research Paper INT-177 43.
McConnell, B. R. 1961. Notes on some rooting characteristics of antelope bitterbrush. PNW Old Series Research Note No. 204:1-5.
McConnell, B. R. and J. G. Smith. 1977. Influence of grazing on age-yield interactions in bitterbrush. Journal of Range Management 30: 91-93.
Merrill, E. H., H. Mayland, and J. Peek. 1982. Shrub responses after fire in an Idaho ponderosa pine community. The Journal of Wildlife Management 46: 496-502.
Miller, R. F. and E. K. Heyerdahl. 2008. Fine-scale variation of historical fire regimes in sagebrush-steppe and juniper woodland: An example from California, USA. International Journal of Wildland Fire 17:245-254.
Miller, R. F. R. J. T. 2000. The role of fire in juniper and pinyon woodlands: A descriptive analysis. Pages p. 15-30 in Proceedings of the Invasive Species Workshop: The Role of Fire in the Control and Spread of Invasive Species. Tallahassee, Florida.
Miller, R. F. C., Jeanne C.; Pyke, David A.; Pierson, Fred B.; Williams, C. Jason 2013. A review of fire effects on vegetation and soils in the Great Basin region: Response and ecological site characteristics. Gen. Tech. Rep. RMRS-GTR-308. Fort Collins CO: U.S. Department of Agriculture, United State Forest Service, Rocky Mountain Research Station. p. 126.
Mueggler, W. F. and J. P. Blaisdell. 1951. Replacing wyethia with desireable forage species. Journal of Range Management 4:143-150.
Murray, R. 1983. Response of antelope bitterbrush to burning and spraying in southeastern Idaho. Tiedemann, Arthur R.; Johnson, Kendall L., compilers. Research and management of bitterbrush and cliffrose in western North America. General Technical Report INT-152. Ogden, UT: US Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: 142-152.
National Oceanic and Atmospheric Administration. 2004. The North American Monsoon. Reports to the Nation. Weather Service, Climate Prediction Center. Available online: http://www.weather.gov/
Neuenschwander, L. 1980. Broadcast burtning of sagebrush in the winter. Journal of Range Management: 233-236.
Noste, N. V. and C. L. Bushey. 1987. Fire response of shrubs of dry forest habitat types in Montana and Idaho. General technical report INT.
Noy-Meir, I. 1973. Desert ecosystems: Environment and producers. Annual Review of ecology and systematics. Vol. 4:25-51.
Personius, T.L., C. L. Wambolt, J. R. Stephens and R. G. Kelsey. 1987. Crude terpenoid influence on mule deer preference for sagebrush. Journal of Range Management, 40:1 p. 84-88.
NRCS Plants Database. (http://plants.usda.gov/)
Richards, J. H. and M. M. Caldwell. 1987. Hydraulic lift: Substantial nocturnal water transport between soil layers by Artemisia tridentata roots. Oecologia 73: 486-489.
Robberecht, R. and G. Defossé. 1995. The relative sensitivity of two bunchgrass species to fire. International Journal of Wildland Fire 5:127-134.
Sheehy, D. P. and A. Winward. 1981. Relative palatability of seven Artemisia taxa to mule deer and sheep. Journal of Range Management 34: 397-399.
Smith, J. K. and W. C. Fischer. 1997. Fire ecology of the forest habitat types of northern Idaho. US Department of Agriculture, Forest Service, Intermountain Research Station.
Tisdale, E. W. and M. Hironaka. 1981. The sagebrush-grass region: A review of the ecological literature. University of Idaho, Forest, Wildlife and Range Experiment Station.
Uresk, D. W., J. F. Cline, and W. H. Rickard. 1976. Impact of wildfire on three perennial grasses in south-central Washington. Journal of Range Management 29: 309-310.
Whisenant, S. G. 1990. Changing fire frequencies on Idaho's Snake River Plains: Ecological and management implications. Pages 4-10 in Symposium on cheatgrass invasion, shrub die-off, and other aspects of shrub biology and management. General Technical Report, Intermountain Research Station, USDA Forest Service.
Wood, M. K., Bruce A. Buchanan, & William Skeet. 1995. Shrub preference and utilization by big game on New Mexico reclaimed mine land. Journal of Range Management 48: 431-437.
Wright, H. A. 1971. Why squirreltail is more tolerant to burning than needle-and-thread. Journal of Range Management 24: 277-284.
Wright, H. A. 1985. Effects of fire on grasses and forbs in sagebrush-grass communities. Pages 12-21 in Rangeland Fire Effects; A Symposium: Boise, ID, USDI-BLM.
Wright, H.A., L.F. Neuenschwander, and C.M. Britton. 1979. The role and use of fire in sagebrush-grass and pinyon-juniper plant communities: a state-of-the-art review. Gen. Tech. Rep. INT-58. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 48 p.
Young, R.P. 1983. Fire as a vegetation management tool in rangelands of the intermountain region. In: Monsen, S.B. and N. Shaw (eds). Managing intermountain rangelands - improvement of range and wildlife habitats: Proceedings of symposia; 1981 September 15-17; Twin Falls, ID; 1982 June 22-24; Elko, NV. Gen. Tech. Rep. INT-157. Ogden, UT. U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. Pgs 18-31.
Ziegenhagen, L. L. 2003. Shrub reestablishment following fire in the mountain big sagebrush (Artemisia tridentata Nutt. ssp. vaseyana (Rydb.) Beetle) alliance. M.S. Oregon State University.
Ziegenhagen, L. L. and R. F. Miller. 2009. Postfire recovery of two shrubs in the interiors of large burns in the intermountain west, USA. Western North American Naturalist 69:195-205.
Zschaechner, G. A. 1985. Studying rangeland fire effects: A case study in Nevada. Pages 66-84 in Rangeland Fire Effects, a Symposium. Bureau of Land Management, Boise, Idaho.
Contributors
RK
TK Stringham
P Novak-Echenique
Approval
Kendra Moseley, 4/25/2024
Rangeland health reference sheet
Interpreting Indicators of Rangeland Health is a qualitative assessment protocol used to determine ecosystem condition based on benchmark characteristics described in the Reference Sheet. A suite of 17 (or more) indicators are typically considered in an assessment. The ecological site(s) representative of an assessment location must be known prior to applying the protocol and must be verified based on soils and climate. Current plant community cannot be used to identify the ecological site.
| Author(s)/participant(s) | Patti Novak-Echenique |
|---|---|
| Contact for lead author | State Rangeland Management Specialist |
| Date | 09/24/2009 |
| Approved by | Kendra Moseley |
| Approval date | |
| Composition (Indicators 10 and 12) based on | Annual Production |
Indicators
-
Number and extent of rills:
Rills are typically none. There may be a few, widely spaced and shallow rills on steeper slopes (30% gradient). -
Presence of water flow patterns:
Water flow patterns are typically non-existent but can rarely occur on steeper slopes (30% gradient) in areas recently subjected to intense summer convection storms or rapid snowmelt. -
Number and height of erosional pedestals or terracettes:
Pedestals are none to rare. -
Bare ground from Ecological Site Description or other studies (rock, litter, lichen, moss, plant canopy are not bare ground):
Bare ground ± 10-20%. -
Number of gullies and erosion associated with gullies:
Gullies are non-existent. -
Extent of wind scoured, blowouts and/or depositional areas:
None -
Amount of litter movement (describe size and distance expected to travel):
Fine litter (foliage from grasses and annual & perennial forbs) expected to move distance of slope length during intense summer convection storms or rapid snowmelt events. Persistent litter (large woody material) will remain in place except during catastrophic events. -
Soil surface (top few mm) resistance to erosion (stability values are averages - most sites will show a range of values):
Soil stability values should be 3 to 6 on most soil textures found on this site. (To be field tested.) -
Soil surface structure and SOM content (include type of structure and A-horizon color and thickness):
Surface structure is fine granular, weak thin platy, or subangular blocky. Soil surface colors are very dark and soils are typified by a mollic epipedon. Organic matter of the surface 2 to 4 inches is typically 1 to 3 percent. Organic matter content can be more or less depending on micro-topography. -
Effect of community phase composition (relative proportion of different functional groups) and spatial distribution on infiltration and runoff:
Perennial herbaceous plants (especially deep-rooted bunchgrasses [i.e. Idaho fescue] slow runoff and increase infiltration. Shrub canopy and associated litter break raindrop impact and provide opportunity for snow catch and accumulation on site. -
Presence and thickness of compaction layer (usually none; describe soil profile features which may be mistaken for compaction on this site):
Compacted layers are not typical. Subsoil subangular blocky structure is not to be interpreted as compacted layers. -
Functional/Structural Groups (list in order of descending dominance by above-ground annual-production or live foliar cover using symbols: >>, >, = to indicate much greater than, greater than, and equal to):
Dominant:
Reference Plant Community: Deep-rooted, cool season, perennial bunchgrassesSub-dominant:
tall shrubs (antelope bitterbrush & mountain big sagebrush) > deep-rooted, cool season, perennial forbs > fibrous, shallow-rooted, cool season, perennial and annual forbs > shallow-rooted, cool season, perennial grasses > associated shrubsOther:
Additional:
-
Amount of plant mortality and decadence (include which functional groups are expected to show mortality or decadence):
Dead branches within individual shrubs not uncommon and standing dead shrub canopy material may be as much as 15% of total woody canopy; some of the mature bunchgrasses (<10%) have dead centers. -
Average percent litter cover (%) and depth ( in):
Between plant interspaces (± 15%) and litter depth is ± 1/4 inch. -
Expected annual annual-production (this is TOTAL above-ground annual-production, not just forage annual-production):
For normal or average growing season (through June) ± 700 lbs/ac. -
Potential invasive (including noxious) species (native and non-native). List species which BOTH characterize degraded states and have the potential to become a dominant or co-dominant species on the ecological site if their future establishment and growth is not actively controlled by management interventions. Species that become dominant for only one to several years (e.g., short-term response to drought or wildfire) are not invasive plants. Note that unlike other indicators, we are describing what is NOT expected in the reference state for the ecological site:
Douglas rabbitbrush is an increaser on this site. Cheatgrass is an invader on this site. -
Perennial plant reproductive capability:
All functional groups should reproduce in average (or normal) and above average growing season years.
Print Options
Sections
Font
Other
The Ecosystem Dynamics Interpretive Tool is an information system framework developed by the USDA-ARS Jornada Experimental Range, USDA Natural Resources Conservation Service, and New Mexico State University.
Click on box and path labels to scroll to the respective text.