
Natural Resources
Conservation Service
Ecological site R025XY057NV
SHALLOW CLAY LOAM 10-14 P.Z.
Last updated: 4/25/2024
Accessed: 04/12/2026
General information
Provisional. A provisional ecological site description has undergone quality control and quality assurance review. It contains a working state and transition model and enough information to identify the ecological site.
MLRA notes
Major Land Resource Area (MLRA): 025X–Owyhee High Plateau
MLRA Notes 25—Owyhee High Plateau
This area is in Nevada (56 percent), Idaho (30 percent), Oregon (12 percent), and Utah (2 percent). It makes up about 27,443 square miles. MLRA 25 is characteristically cooler and wetter than the neighboring MLRAs of the Great Basin. The western boundary is marked by a gradual transition to the lower and warmer basins of MLRA 24. The boundary to the south-southeast, with MLRA 28B, is marked by gradual changes in geology marked by an increased dominance of singleleaf pinyon and Utah juniper and a reduced presence of Idaho fescue. The boundary to the north, with MLRA 11, is a rapid transition from the lava plateau topography to the lower elevation Snake River Plain.
Physiography:
All of this area lies within the Intermontane Plateaus. The southern half is in the Great Basin section of the Basin and Range province. This part of the MLRA is characterized by isolated, uplifted fault-block mountain ranges separated by narrow, aggraded desert plains. This geologically older terrain has been dissected by numerous streams draining to the Humboldt River.
The northern half of the area lies within the Columbia Plateaus province. This part of the MLRA forms the southern boundary of the extensive Columbia Plateau basalt flows. Most of the northern half is in the Payette section, but the northeast corner is in the Snake River Plain section. Deep, narrow canyons draining into the Snake River have been incised into this broad basalt plain. Elevation ranges from 3,000 to 7,550 feet on rolling plateaus and in gently sloping basins. It is more than 9,840 feet on some steep mountains. The Humboldt River crosses the southern half of this area
Geology:
The dominant rock types in this MLRA are volcanic. They include andesite, basalt, tuff, and rhyolite. In the north and west parts of the area, Cretaceous granitic rocks are exposed among Miocene volcanic rocks in mountains. A Mesozoic igneous and metamorphic rock complex dominates the south and east parts of the area. Upper and Lower Paleozoic calcareous sediments, including oceanic deposits, are exposed with limited extent in the mountains. Alluvial fan and basin fill sediments occur in the valleys.
Climate:
The average annual precipitation in most of this area is typically 11 to 22 inches. It increases to as much as 49 inches at the higher elevations. Rainfall occurs in spring and sporadically in summer. Precipitation occurs mainly as snow in winter. The precipitation is distributed fairly evenly throughout fall, winter, and spring. The amount of precipitation is lowest from midsummer to early autumn. The average annual temperature is 33 to 51 degrees F. The freeze-free period averages 130 days and ranges from 65 to 190 days, decreasing in length with elevation. It is typically less than 70 days in the mountains.
Water:
The supply of water from precipitation and streamflow is small and unreliable, except along the Owyhee, Bruneau, and Humboldt Rivers. Streamflow depends largely on accumulated snow in the mountains. Surface water from mountain runoff is generally of excellent quality and suitable for all uses. The basin fill sediments in the narrow alluvial valleys between the mountain ranges provide some ground water for irrigation. The alluvial deposits along the large streams have the most ground water. Based on measurements of water quality in similar deposits in adjacent areas, the basin fill deposits probably contain moderately hard water. The water is suitable for almost all uses. The carbonate rocks in this area are considered aquifers, but they are little used. Springs are common along the edges of the limestone outcrops.
Soils:
The dominant soil orders in this MLRA are Aridisols and Mollisols. The soils in the area dominantly have a mesic or frigid temperature regime and an aridic, aridic bordering on xeric, or xeric moisture regime. Soils with aquic moisture regimes are limited to drainage or spring areas, where moisture originates or runs on and through. These soils are of a very limited extent throughout the MLRA. They generally are well drained, clayey or loamy, and shallow or moderately deep. Most of the soils formed in mixed parent material. Volcanic ash and loess mantle the landscape. Surface soil textures are loam and silt loam with ashy texture modifiers in some areas. Argillic horizons occur on the more stable landforms. They are exposed nearer the soil surface on convex landforms, where ash and loess deposits are more likely to erode. Soils that formed in carbonatic parent material in areas that receive less than 12 inches of precipitation are characterized by calcic horizons throughout the profile, while soils in areas that receive more than 12 inches of precipitation do not have calcic horizons in the upper part of the profile. Soils that formed on stable landforms at the lower elevations are dominated by ochric horizons. Soils that formed at the middle and upper elevations are characterized by mollic epipedons. Soils in drainage areas at all elevations that receive moisture running on or through them are characterized by thicker mollic epipedons.
Biological Resources:
This MLRA supports shrub-grass vegetation. Lower elevations are characterized by Wyoming big sagebrush associated with bluebunch wheatgrass, western wheatgrass, and Thurber’s needlegrass. Other important plants include bluegrass, squirreltail, penstemon, phlox, milkvetch, lupine, Indian paintbrush, aster, and rabbitbrush. Black sagebrush occurs but is less extensive. Singleleaf pinyon and Utah juniper occur in limited areas. With increasing elevation and precipitation, vast areas characterized by mountain big sagebrush or low sagebrush/early sagebrush in association with Idaho fescue, bluebunch wheatgrass, needlegrasses, and bluegrass become common. Snowberry, curl-leaf mountain mahogany, ceanothus, and juniper also occur. Mountains at the highest elevations support whitebark pine, Douglas-fir, limber pine, Engelmann spruce, subalpine fir, aspen, and curl-leaf mountain mahogany.
Major wildlife species include mule deer, bighorn sheep, pronghorn, mountain lion, coyote, bobcat, badger, river otter, mink, weasel, golden eagle, red-tailed hawk, ferruginous hawk, Swainson’s hawk, northern harrier, prairie falcon, kestrel, great horned owl, short-eared owl, long-eared owl, burrowing owl, pheasant, sage grouse, chukar, gray partridge, and California quail. Reptiles and amphibians include western racer, gopher snake, western rattlesnake, side-blotched lizard, western toad, and spotted frog. Fish species include bull, red band, and rainbow trout.
Ecological site concept
This site occurs on summits and upper backslopes of hills and lower mountains on all aspects. Slopes range from 4 to 75 percent, but slope gradients of 15 to 50 percent are most typical. Elevations are 5500 to 8500 feet. The average growing season is about 80 to 100 days.
The soils associated with this site are shallow to very shallow and well drained. Textures are sandy clay loams and coarse sandy loams, modified by 15 to 30 percent rock fragments. The soils typically have a mollic epipedon and an argillic horizon. The soil moisture regime is xeric bordering on aridic. The soil temperature regime is frigid.
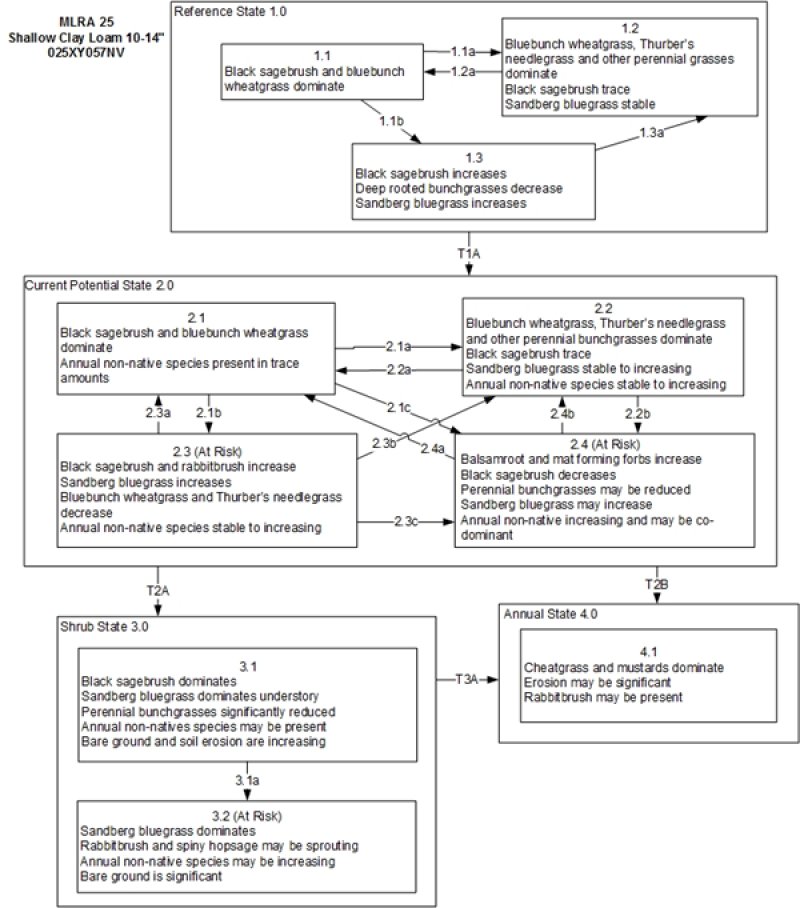
The reference plant community is dominated by Thurber''s needlegrass, bluebunch wheatgrass and black sagebrush. Potential vegetative composition is about 55% grasses, 10% forbs and 35% shrubs. Approximate ground cover (basal and crown) is 15 to 30 percent.
Associated sites
| R025XY009NV |
SOUTH SLOPE 12-14 P.Z. |
|---|---|
| R025XY014NV |
LOAMY 10-12 P.Z. |
| R025XY058NV |
BOULDERY LOAM |
Similar sites
| R025XY026NV |
CHANNERY HILL ACHY dominant grass; less productive site |
|---|---|
| R025XY025NV |
CHALKY KNOLL ARTRW8 codominant shrub; ACHY-ELEL5 codominant grasses; less productive site |
| R025XY055NV |
SHALLOW CLAY SLOPE 10-14 P.Z. PSSPS dominant grass; less productive site; steeper slopes |
| R025XY024NV |
MOUNTAIN RIDGE Higher elevations; FEID dominant grass |
Table 1. Dominant plant species
| Tree |
Not specified |
|---|---|
| Shrub |
(1) Artemisia nova |
| Herbaceous |
(1) Pseudoroegneria spicata subsp. spicata |






Click on box and path labels to scroll to the respective text.