Natural Resources
Conservation Service
Ecological site R028AY022NV
GRAVELLY CLAY 8-10 P.Z.
Accessed: 04/14/2026
General information
Provisional. A provisional ecological site description has undergone quality control and quality assurance review. It contains a working state and transition model and enough information to identify the ecological site.
Figure 1. Mapped extent
Areas shown in blue indicate the maximum mapped extent of this ecological site. Other ecological sites likely occur within the highlighted areas. It is also possible for this ecological site to occur outside of highlighted areas if detailed soil survey has not been completed or recently updated.
MLRA notes
Major Land Resource Area (MLRA): 028A–Ancient Lake Bonneville
MLRA 28A occurs in Utah (82%), Nevada (16%), and Idaho (2%). It makes up about 36,775 square miles. A large area west and southwest of Great Salt Lake is a salty playa. This area is the farthest eastern extent of the Great Basin Section of the Basin and Range Province of the Intermontane Plateaus. It is an area of nearly level basins between widely separated mountain ranges trending north to south. The basins are bordered by long, gently sloping alluvial fans. The mountains are uplifted fault blocks with steep side slopes. They are not well dissected because of low rainfall in the MLRA. Most of the valleys are closed basins containing sinks or playa lakes. Elevation ranges from 3,950 to 6,560 ft. in the basins and from 6,560 to 11,150 ft. in the mountains. Most of this area has alluvial valley fill and playa lakebed deposits at the surface. Great Salt Lake is all that remains of glacial Lake Bonneville. A level line on some mountain slopes indicates the former extent of this glacial lake. Most of the mountains in the interior of this area consist of tilted blocks of marine sediments from Cambrian to Mississippian age. Scattered outcrops of Tertiary continental sediments and volcanic rocks are throughout the area. The average annual precipitation is 5 to 12 ins. in the valleys and is as much as 49 ins. in the mountains. Most of the rainfall occurs as high-intensity, convective thunderstorms during the growing season. The driest period is from midsummer to early autumn. Precipitation in winter typically occurs as snow. The average annual temperature is 39 to 53 °F. The freeze-free period averages 165 days and ranges from 110 to 215 days, decreasing in length with elevation. The dominant soil orders in this MLRA are Aridisols, Entisols, and Mollisols. The soils in the area dominantly have a mesic or frigid soil temperature regime, an aridic or xeric soil moisture regime, and mixed mineralogy. They generally are well drained, loamy or loamy-skeletal, and very deep.
Ecological site concept
This site occurs on low mountains and rolling hills on all aspects. Slopes range from 8 to 30 percent, but slope gradients of 8 to 15 percent are typical. Elevations are 6000 to 7000 feet.
Average annual precipitation is 8 to 10(11) inches. Mean annual air temperature is 45 to 50 degrees F. The average growing season is about 100 to 120 days.
Soils associated with this site are moderately deep and well drained. Surface soils are medium to coarse textured and subsoils are medium to fine textured.
The reference state is dominated by Thurber's needlegrass and Wyoming big sagebrush. Production ranges from 350 to 800 pounds per acre.
Associated sites
| R028AY013NV |
SHALLOW CALCAREOUS LOAM 8-10 P.Z. |
|---|---|
| R028AY015NV |
LOAMY 8-10 P.Z. |
| R028AY017NV |
SHALLOW LOAM 8-10 P.Z. |
| R028AY050NV |
GRAVELLY CLAY 10-12 P.Z. |
Similar sites
| R028AY005NV |
SANDY 8-10 P.Z. ACTH7 absent |
|---|---|
| R028AY015NV |
LOAMY 8-10 P.Z. ACTH7 absent |
| R028AY028NV |
DROUGHTY LOAM 8-10 P.Z. ACTH7 absent |
| R028AY031NV |
LOAMY FAN 8-10 P.Z. LECI4-ELLAL codominant |
| R028AY040NV |
GRAVELLY LOAM 10-12 P.Z. ACTH7 absent; PUST codominant shrub |
| R028AY050NV |
GRAVELLY CLAY 10-12 P.Z. PSSPS dominant grass |
| R028AY054NV |
COARSE LOAMY FAN 8-10 P.Z. LECI4-ACHY codominant; ACTH7 absent |
| R028AY017NV |
SHALLOW LOAM 8-10 P.Z. ACTH7 absent |
| R028AY010NV |
COARSE GRAVELLY LOAM 10-12 P.Z. ACTH7 absent |
Table 1. Dominant plant species
| Tree |
Not specified |
|---|---|
| Shrub |
(1) Artemisia tridentata ssp. wyomingensis |
| Herbaceous |
(1) Achnatherum thurberianum |
Physiographic features
This site occurs on low mountains and rolling hills on all aspects. Slopes range from 8 to 30 percent, but slope gradients of 8 to 15 percent are typical. Elevations are 6000 to 7000 feet.
Table 2. Representative physiographic features
| Landforms |
(1)
Mountain
(2) Hill |
|---|---|
| Elevation | 6,000 – 7,000 ft |
| Slope | 8 – 30% |
| Aspect | Aspect is not a significant factor |
Climatic features
Nevada’s climate is predominantly arid, with large daily ranges of temperature, infrequent severe storms, heavy snowfall in the higher mountains, and great location variations with elevation. Three basic geographical factors largely influence Nevada’s climate: continentality, latitude, and elevation. Continentality is the most important factor. The strong continental effect is expressed in the form of both dryness and large temperature variations. Nevada lies on the eastern, lee side of the Sierra Nevada Range, a massive mountain barrier that markedly influences the climate of the State. The prevailing winds are from the west, and as the warm moist air from the Pacific Ocean ascend the western slopes of the Sierra Range, the air cools, condensation occurs and most of the moisture falls as precipitation. As the air descends the eastern slope, it is warmed by compression, and very little precipitation occurs. The effects of this mountain barrier are felt not only in the West but throughout the state, with the result that the lowlands of Nevada are largely desert or steppes. The temperature regime is also affected by the blocking of the inland-moving maritime air. Nevada sheltered from maritime winds, has a continental climate with well-developed seasons and the terrain responds quickly to changes in solar heating.
Nevada lies within the mid-latitude belt of prevailing westerly winds which occur most of the year. These winds bring frequent changes in weather during the late fall, winter and spring months, when most of the precipitation occurs. To the south of the mid-latitude westerlies, lies a zone of high pressure in subtropical latitudes, with a center over the Pacific Ocean. In the summer, this high-pressure belt shifts northward over the latitudes of Nevada, blocking storms from the ocean. The resulting weather is mostly clear and dry during the summer and early fall, with scattered thundershowers. The eastern portion of the state receives significant summer thunderstorms generated from monsoonal moisture pushed up from the Gulf of California, known as the North American monsoon. The monsoon system peaks in August and by October the monsoon high over the Western U.S. begins to weaken and the precipitation retreats southward towards the tropics (NOAA 2004).
Average annual precipitation is 8 to 10(11) inches. Mean annual air temperature is 45 to 50 degrees F. The average growing season is about 100 to 120 days.
Mean annual precipitation at the LUND, NEVADA climate station (264745) is 9.61 inches.
Monthly mean precipitation is:
January 0.78; February 0.85; March 1;
April 0.98; May 0.95; June 0.82; July 0.69;
August 0.87; September 0.77; October 0.92;
November 0.69; December 0.73.
Table 3. Representative climatic features
| Frost-free period (average) | 0 days |
|---|---|
| Freeze-free period (average) | 110 days |
| Precipitation total (average) | 9 in |
Figure 2. Monthly average minimum and maximum temperature
Figure 3. Annual precipitation pattern
Figure 4. Annual average temperature pattern
Influencing water features
There are no influencing water features associated with this site.
Soil features
Soils associated with this site are moderately deep and well drained. Surface soils are medium to coarse textured and subsoils are medium to fine textured. The available water holding capacity is very low. Some soils are modified with high volumes of rock fragments throughout the soil profile. Runoff is medium and the potential for sheet and rill erosion varies with slope gradient.
The representative soil series is Maderbak, a clayey-skeletal, smectitic, mesic Xeric Haplargid. The soils have an ochric epipedon from the surface to 3 inches and an argillic horizon from 3 to 17 inches. Clay content in the particle-size control section ranges from 35 to 50 percent. Reaction is slightly to moderately alkaline. Effervescence is violently effervescent below 17 inches. Lithology consists of volcanic rocks.
Table 4. Representative soil features
| Parent material |
(1)
Colluvium
–
andesite
(2) Residuum – andesite |
|---|---|
| Surface texture |
(1) Very gravelly clay loam |
| Family particle size |
(1) Clayey |
| Drainage class | Well drained |
| Permeability class | Moderately rapid |
| Soil depth | 20 – 40 in |
| Surface fragment cover <=3" | 25 – 40% |
| Surface fragment cover >3" | 5 – 30% |
| Available water capacity (0-40in) |
2.6 – 2.7 in |
| Calcium carbonate equivalent (0-40in) |
Not specified |
| Electrical conductivity (0-40in) |
Not specified |
| Sodium adsorption ratio (0-40in) |
Not specified |
| Soil reaction (1:1 water) (0-40in) |
7.8 – 8.2 |
| Subsurface fragment volume <=3" (Depth not specified) |
35 – 55% |
| Subsurface fragment volume >3" (Depth not specified) |
5% |
Ecological dynamics
An ecological site is the product of all the environmental factors responsible for its development and it has a set of key characteristics that influence a site’s resilience to disturbance and resistance to invasion. Key characteristics include 1) climate (precipitation, temperature), 2) topography (aspect, slope, elevation, and landform), 3) hydrology (infiltration, runoff), 4) soils (depth, texture, structure, organic matter), 5) plant communities (functional groups, productivity), and 6) natural disturbance regime (fire, herbivory, etc.) (Caudle et al. 2013). Biotic factors that influence resilience include site productivity, species composition and structure, and population regulation and regeneration (Chambers et al. 2013).
This ecological site is dominated by deep-rooted cool season, perennial bunchgrasses and long-lived shrubs (50+ years) with high root to shoot ratios. The dominant shrubs usually root to the full depth of the winter-spring soil moisture recharge, which ranges from 1.0 to over 3.0 m. (Comstock and Ehleringer 1992). Root length of mature sagebrush plants was measured to a depth of 2 meters in alluvial soils in Utah (Richards and Caldwell 1987). These shrubs have a flexible generalized root system with development of both deep taproots and laterals near the surface (Dobrowolski et al. 1990).
In the Great Basin, the majority of annual precipitation is received during the winter and early spring. This continental semiarid climate regime favors growth and development of deep-rooted shrubs and herbaceous cool season plants using the C3 photosynthetic pathway (Comstock and Ehleringer 1992). Winter precipitation and slow melting of snow results in deeper percolation of moisture into the soil profile. Herbaceous plants, more shallow-rooted than shrubs, grow earlier in the growing season and thrive on spring rains, while the deeper rooted shrubs lag in phenological development because they draw from deeply infiltrating moisture from snowmelt the previous winter. Periodic drought regularly influences sagebrush ecosystems and drought duration and severity has increased throughout the 20th century in much of the Intermountain West. Major shifts away from historical precipitation patterns have the greatest potential to alter ecosystem function and productivity. Species composition and productivity can be altered by the timing of precipitation and water availability within the soil profile (Bates et al 2006).
Variability in plant community composition and production depends on soil surface texture and depth. Needleandthread grass is adapted to coarser textured soils whereas Indian ricegrass will increase with sandy soil surfaces, and bottlebrush squirreltail will increase with silty soil surfaces. Production generally increases with soil depth. The amount of sagebrush in the plant community is dependent upon disturbances like fire, Aroga moth infestations, and grazing.
Wyoming big sagebrush, the most drought tolerant of the big sagebrushes, is generally long-lived; therefore it is not necessary for new individuals to recruit every year for perpetuation of the stand. Infrequent large recruitment events and simultaneous low, continuous recruitment is the foundation of population maintenance (Noy-Meir 1973). Survival of the seedlings is dependent on adequate moisture conditions.
Native insect outbreaks are also important drivers of ecosystem dynamics in sagebrush communities. Climate is generally believed to influence the timing of insect outbreaks especially a sagebrush defoliator, Aroga moth (Aroga websteri). Aroga moth infestations have occurred in the Great Basin in the 1960s, early 1970s, and is ongoing in Nevada since 2004 (Bentz, et al 2008). Thousands of acres of big sagebrush have been impacted, with partial to complete die-off observed. Aroga moth can partially or entirely kill individual plants or entire stands of big sagebrush (Furniss and Barr 1975).
Perennial bunchgrasses generally have somewhat shallower root systems than shrubs in these systems, but root densities are often as high as or higher than those of shrubs in the upper 0.5 m but taper off more rapidly than shrubs. General differences in root depth distributions between grasses and shrubs result in resource partitioning in these shrub/grass systems.
The Great Basin sagebrush communities have high spatial and temporal variability in precipitation, both among years and within growing seasons. Nutrient availability is typically low but increases with elevation and closely follows moisture availability. The moisture resource supporting the greatest amount of plant growth is usually the water stored in the soil profile during the winter. The invasibility of plant communities is often linked to resource availability. Disturbance can decrease resource uptake due to damage or mortality of the native species and depressed competition or can increase resource pools by the decomposition of dead plant material following disturbance. The invasion of sagebrush communities by cheatgrass has been linked to disturbances (fire, abusive grazing) that have resulted in fluctuations in resources (Chambers et al. 2007).
The introduction of annual weedy species, like cheatgrass, may cause an increase in fire frequency and eventually lead to an annual state. Conversely, as fire frequency decreases, sagebrush will increase and with inappropriate grazing management the perennial bunchgrasses and forbs may be reduced.
At the upper range of this site's precipitation range, there is potential for infilling by singleleaf pinyon and Utah juniper. Infilling may also occur if the site is adjacent to woodland sites or other ecological sites with pinyon and juniper present. Without disturbance in these areas, singleleaf pinyon and Utah juniper will eventually dominate the site and out-compete sagebrush for water and sunlight severely reducing both the shrub and herbaceous understory (Miller and Tausch 2000, Lett and Knapp 2005). The potential for soil erosion increases as the woodland matures and the understory plant community cover declines (Pierson et al. 2010).
This ecological site has low resilience to disturbance and low resistance to invasion. Resilience increases with elevation, aspect, increased precipitation and increased nutrient availability. Five possible alternative stable states have been identified for this site.
Fire Ecology:
Wyoming big sagebrush communities historically had low fuel loads, and patchy fires that burned in a mosaic pattern were common at 10 to 70 year return intervals (Young et al. 1979, West and Hassan 1985, Bunting et al. 1987). Davies et al. (2006) suggest historic fire return intervals in Wyoming big sagebrush communities were around 50 to 100 years. More recently, Baker (2011) estimates fire rotation to be 200 to 350 years in Wyoming big sagebrush communities. Wyoming big sagebrush is killed by fire and only regenerates from seed. Recovery time for Wyoming big sagebrush may require 50 to 120 or more years (Baker 2006). However, the introduction and expansion of cheatgrass has dramatically altered the fire regime (Balch et al. 2013) and restoration potential of Wyoming big sagebrush communities.
The effect of fire on bunchgrasses relates to culm density, culm-leaf morphology, and the size of the plant. The initial condition of bunchgrasses within the site along with seasonality and intensity of the fire all factor into the individual species response. For most forbs and grasses the growing points are located at or below the soil surface providing relative protection from disturbances which decrease above ground biomass, such as grazing or fire. Thus, fire mortality is more correlated to duration and intensity of heat which is related to culm density, culm-leaf morphology, size of plant and abundance of old growth (Young 1983, Wright 1971).
Burning has been found to decrease the vegetative and reproductive vigor of Thurber’s needlegrass (Uresk et al. 1976). Fire can cause high mortality, in addition to reducing basal area and yield of Thurber’s needlegrass (Britton et al. 1990). The fine leaves and densely tufted growth form make this grass susceptible to subsurface charring of the crowns (Wright and Klemmedson 1965). Although timing of fire highly influenced the response and mortality of Thurber’s needlegrass, smaller bunch sizes were less likely to be damaged by fire (Wright and Klemmedson 1965). Thurber’s needlegrass often survives fire and will continue growth or regenerate from tillers when conditions are favorable (Koniak 1985, Britton et al. 1990). Reestablishment on burned sites has been found to be relatively slow due to low germination and competitive ability (Koniak 1985). Cheatgrass has been found to be a highly successful competitor with seedlings of this needlegrass and may preclude reestablishment (Evans and Young 1978)Indian ricegrass is fairly fire tolerant (Wright 1985), which is likely due to its low culm density and below ground plant crowns. Indian ricegrass has been found to reestablish on burned sites through seed dispersed from adjacent unburned areas (Young 1983, West 1994). Thus the presence of surviving, seed producing plants is necessary for reestablishment of Indian ricegrass. Grazing management following fire to promote seed production and establishment of seedlings is important.
Needleandthread is top-killed by fire but is likely to resprout if fire does not consume above ground stems (Akinsoji 1988, Bradley, Noste and Fischer 1992). In a study by Wright and Klemmedson (1965), season of burn rather than fire intensity seemed to be the crucial factor in mortality for needle-and-thread grass. Early spring season burning was seen to kill the plants while August burning had no effect. Needleandthread is a fine leaf grass and is considered sensitive to fire (Akinsoji 1988, Bradley et al. 1992, Miller et al. 2013). In a study by Wright and Klemmedson (1965), season of burn rather than fire intensity seemed to be the crucial factor in mortality for needle-and-thread grass. Early spring season burning was seen to kill the plants while August burning had no effect. Thus, under wildfire scenarios needle-and-thread is often present in the post-burn community.
Galleta grass, a minor component of these ecological sites, has been found to increase following fire likely due to its rhizomatous root structure and ability to resprout (Jameson 1962). Galleta is found more often in the southern section of MLRA 28A, where monsoonal rains are more common. Sandberg bluegrass, another minor component of these ecological sites, has also been found to increase following fire likely due to its low stature and productivity (Daubenmire 1975). Both grass species may retard reestablishment of deeper rooted bunchgrasses. Repeated frequent fire in this community will eliminate Wyoming sagebrush, significantly decrease bunchgrass density on the site and facilitate the establishment of an annual weed community with varying amounts of galleta, Sandberg bluegrass, spiny hopsage, and rabbitbrush.
The range and density of singleleaf pinyon and Utah juniper has increased since the middle of the nineteenth century (Tausch 1999, Miller and Tausch 2000). Causes for expansion of trees into sagebrush ecosystems include wildfire suppression, historic livestock grazing, and climate change (Bunting 1994).
State and transition model
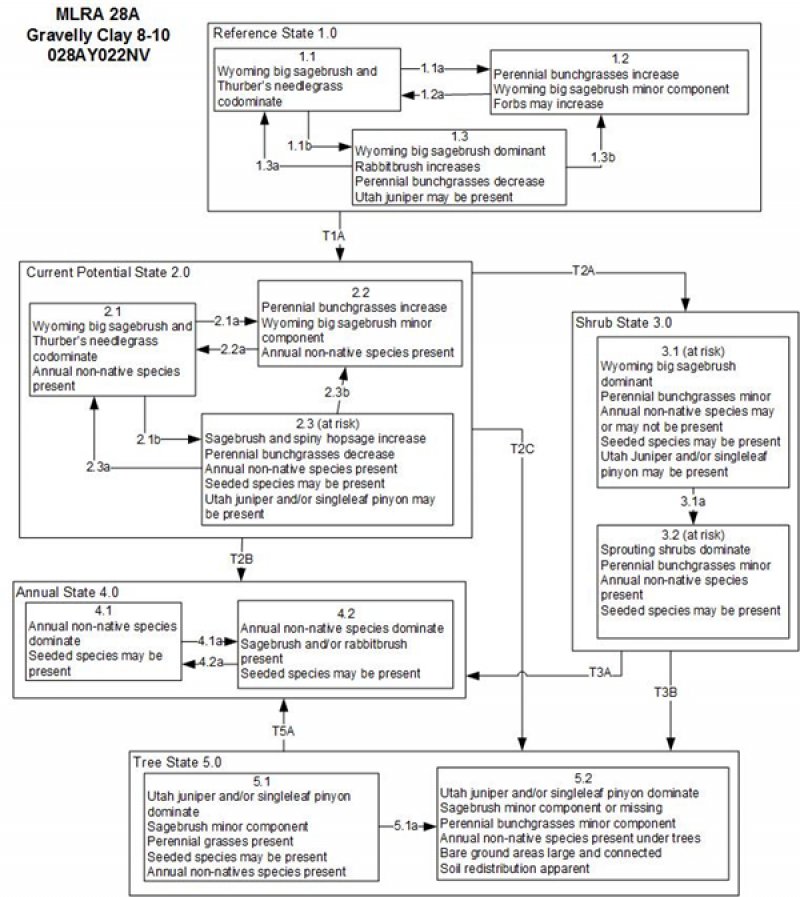
Figure 5. State and Transition Model

Figure 6. Legend
More interactive model formats are also available.
View Interactive Models
More interactive model formats are also available.
View Interactive Models
Click on state and transition labels to scroll to the respective text
Ecosystem states
States 2 and 5 (additional transitions)
State 1 submodel, plant communities
State 2 submodel, plant communities
State 3 submodel, plant communities
State 4 submodel, plant communities
State 5 submodel, plant communities
State 1
Reference State
The Reference State 1.0 is a representative of the natural range of variability under pristine conditions. The reference state has 3 general community phases; a shrub-grass dominant phase, a perennial grass dominant phase and a shrub dominant phase. State dynamics are maintained by interactions between climatic patterns and disturbance regimes. Negative feedbacks enhance ecosystem resilience and contribute to the stability of the state. These include the presence of all structural and functional groups, low fine fuel loads, and retention of organic matter and nutrients. Plant community phase changes are primarily driven by fire, periodic drought and/or insect or disease attack.
Community 1.1
Community Phase
Wyoming big sagebrush and Thurber's needlegrass dominate this phase. Indian ricegrass and needleanthread grass are sub-dominants in the understory. Other shrubs include antelope bitterbrush, Nevada ephedra and rabbitbrush. Sandberg bluegrass, galleta grass, squirreltail, and forbs are also common in this phase. Utah juniper may be present. Potential vegetative composition is about 55% grasses, 10% forbs and 35% shrubs and trees. Approximate ground cover (basal and crown) is 25 to 35 percent.
Figure 7. Annual production by plant type (representative values) or group (midpoint values)
Table 5. Annual production by plant type
| Plant type | Low (lb/acre) |
Representative value (lb/acre) |
High (lb/acre) |
|---|---|---|---|
| Grass/Grasslike | 193 | 330 | 440 |
| Shrub/Vine | 116 | 198 | 264 |
| Forb | 35 | 180 | 240 |
| Tree | 7 | 12 | 16 |
| Total | 351 | 720 | 960 |
Community 1.2
Community Phase
This community phase is characteristic of a post-disturbance, early seral community phase. Thurber's needlegrass, Indian ricegrass, needleandthread and other perennial grasses dominate. Wyoming big sagebrush is killed by fire, therefore decreasing within the burned community. Spiny hopsage may resprout. Sagebrush could still be present in unburned patches. Forbs may increase post-fire but will likely return to pre-burn levels within a few years.
Community 1.3
Community Phase
Wyoming sagebrush increases in the absence of disturbance or with grazing management that favors shrubs. Decadent sagebrush dominates the overstory and the deep-rooted perennial bunchgrasses in the understory are reduced either from competition with shrubs or from grazing management. Utah juniper may be present.
Pathway a
Community 1.1 to 1.2
Fire would decrease or eliminate the overstory of sagebrush and allow for the perennial bunchgrasses to dominate the site. Fires would typically be small and patchy due to low fuel loads. A fire following an unusually wet spring or a change in management may be more severe and reduce sagebrush cover to trace amounts. A severe infestation of Aroga moth could also cause a large decrease in sagebrush within the community, giving a competitive advantage to the perennial grasses and forbs.
Pathway b
Community 1.1 to 1.3
Chronic drought, time and/or inappropriate grazing management favor an increase in Wyoming sagebrush over deep-rooted perennial bunchgrasses. Combinations of these would allow the sagebrush overstory to increase and dominate the site, causing a reduction in the perennial bunchgrasses. Sandberg bluegrass may increase in density depending on the grazing management.
Pathway a
Community 1.2 to 1.1
Absence of disturbance and/or grazing management that favors shrubs over time would allow the sagebrush to recover.
Pathway a
Community 1.3 to 1.1
A change in grazing management that decreases shrubs would allow for the perennial bunchgrasses in the understory to increase. Heavy late-fall grazing by sheep will reduce sagebrush and increase the herbaceous understory. A moderate Aroga moth infestation may also reduce sagebrush overstory and allow perennial bunchgrasses to increase.
Pathway b
Community 1.3 to 1.2
Fire would decrease or eliminate the overstory of sagebrush and allow for the perennial bunchgrasses to dominate the site. Fires would typically be small and patchy due to low fuel loads. A fire following an unusually wet spring or a change in management may be more severe and reduce sagebrush cover to trace amounts. A severe infestation of Aroga moth could also cause a large decrease in sagebrush within the community, giving a competitive advantage to the perennial grasses and forbs.
State 2
Current Potential State
This state is similar to the Reference State 1.0. Ecological function has not changed, however the resiliency of the state has been reduced by the presence of invasive weeds. This state has the same three general community phases. Negative feedbacks enhance ecosystem resilience and contribute to the stability of the state. These include the presence of all structural and functional groups, low fine fuel loads and retention of organic matter and nutrients. Positive feedbacks decrease ecosystem resilience and stability of the state. These include the non-natives high seed output, persistent seed bank, rapid growth rate, ability to cross pollinate, and adaptations for seed dispersal. Additionally, the presence of highly flammable, non-native species reduces State resilience because these species can promote fire where historically fire has been infrequent, leading to positive feedbacks that further the degradation of the system.
Community 2.1
Community Phase
Wyoming big sagebrush and Thurber's needlegrass dominate this phase. Indian ricegrass, needleandthread are other common grasses. Sandberg’s bluegrass, galleta, blue grama and forbs make up smaller percentages by weight of the understory. Non-native annual species are present.Utah juniper may be present.
Community 2.2
Community Phase
This community phase is characteristic of a post-disturbance, early seral community phase. Indian ricegrass and other perennial grasses dominate. Wyoming big sagebrush is killed by fire, therefore decreasing within the burned community. Spiny hopsage may resprout. Sagebrush could still be present in unburned patches. Forbs may increase post-fire but will likely return to pre-burn levels within a few years. Annual non-native species generally respond well after fire and may be stable or increasing within the community. Rabbitbrush may dominate the aspect for a number of years following fire.
Community 2.3
Community Phase (at risk)
Wyoming big sagebrush increases and the perennial understory are reduced. Decadent sagebrush dominates the overstory and the deep-rooted perennial bunchgrasses in the understory are reduced either from competition with shrubs or from grazing management. Sandberg bluegrass will likely increase in the understory and may be the dominant grass on the site. Utah juniper may be present. Annual non-natives are present.
Pathway a
Community 2.1 to 2.2
Fire would decrease or eliminate the overstory of sagebrush and allow for the perennial bunchgrasses to dominate the site. Fires would typically be small and patchy due to low fuel loads. A fire following an unusually wet spring or a change in management may be more severe and reduce sagebrush cover to trace amounts. A severe infestation of Aroga moth could also cause a large decrease in sagebrush within the community, giving a competitive advantage to the perennial grasses and forbs. Annual non-native species generally respond well after fire and may be stable or increasing within the community.
Pathway b
Community 2.1 to 2.3
Time, chronic drought, grazing management that favors shrubs or combinations of these would allow the sagebrush overstory to increase and dominate the site, causing a reduction in the perennial bunchgrasses. However, Sandberg bluegrass and/or squirreltail may increase in the understory depending on the grazing management. Heavy spring grazing will favor an increase in sagebrush. Annual non-native species may be stable or increasing within the understory.
Pathway a
Community 2.2 to 2.1
Absence of disturbance over time allows for the sagebrush to recover.
Pathway a
Community 2.3 to 2.1
Aroga moth infestation, release from growing season herbivory or drought or combinations of these allows for perennial bunchgrasses to increase.
Pathway b
Community 2.3 to 2.2
Fire would decrease or eliminate the overstory of sagebrush and allow for the perennial bunchgrasses to dominate the site. Fires would typically be small and patchy due to low fuel loads. A fire following an unusually wet spring or a change in management may be more severe and reduce sagebrush cover to trace amounts. A severe infestation of Aroga moth could also cause a large decrease in sagebrush within the community, giving a competitive advantage to the perennial grasses and forbs.
State 3
Sagebrush State
This state is a product of many years of heavy grazing during time periods harmful to perennial bunchgrasses. Sandberg bluegrass will increase with a reduction in deep rooted perennial bunchgrass competition and become the dominant grass. Sagebrush dominates the overstory and rabbitbrush may be a significant component. Sagebrush cover exceeds site concept and may be decadent, reflecting stand maturity and lack of seedling establishment due to competition with mature plants. The shrub overstory and Sandberg bluegrass understory dominate site resources such that soil water, nutrient capture, nutrient cycling, and soil organic matter are temporally and spatially redistributed. Restoration from this site is unlikely; brush treatments have been seen to fail and instead push the site to an annual state.
Community 3.1
Community Phase (at risk)
Wyoming big sagebrush dominates overstory and rabbitbrush may be a significant component. Perennial bunchgrasses are a minor component. Utah juniper may be present or increasing. Annual non-native species are present to increasing. Understory may be sparse, with bare ground increasing.
Community 3.2
Community Phase (at risk)
Sprouting shrubs such as rabbitbrush or spiny hopsage may dominate aspect following fire for a number of years. Trace amounts of sagebrush may be present.
Pathway a
Community 3.1 to 3.2
Fire would decrease or eliminate the overstory of sagebrush. A severe infestation of Aroga moth could also cause a large decrease in sagebrush within the community, giving a competitive advantage to the Sandberg bluegrass, forbs and sprouting shrubs.
State 4
Annual State
This community is characterized by the dominance of annual non-native species such as cheatgrass and tansy mustard in the understory. Wyoming big sagebrush and/or sprouting shrubs may dominate the overstory.
Community 4.1
Community Phase
Annual non-native plants such as cheatgrass or tansy mustard dominate the site. This phase may have seeded species present if resulting from a failed seeding attempt.
Community 4.2
Community Phase
Wyoming big sagebrush and/or spiny hopsage remain in the overstory with annual non-native species, likely cheatgrass, dominating the understory. Trace amounts of desirable bunchgrasses may be present.
Pathway a
Community 4.1 to 4.2
Time and lack of fire allows for shrubs to reestablish. Sprouting shrubs such as spiny hopsage and rabbitbrush will be the first to reappear after fire. Probability of sagebrush establishment is extremely low.
Pathway a
Community 4.2 to 4.1
Fire removes sagebrush and allows for annual non-native species to dominate the site.
State 5
Tree State
This state is characterized by a dominance of Utah juniper and/or singleleaf pinyon in the overstory. Wyoming big sagebrush and perennial bunchgrasses may still be present, but they are no longer controlling site resources. Soil moisture, soil nutrients and soil organic matter distribution and cycling have been spatially and temporally altered.
Community 5.1
Community Phase
Utah juniper and/or singleleaf pinyon trees dominate overstory, sagebrush is decadent and dying, deep rooted perennial bunchgrasses are decreasing. Recruitment of sagebrush cohorts is minimal. Annual non-natives may be present or increasing.
Community 5.2
Community Phase (at risk)
Utah juniper and/or singleleaf pinyon dominate the site and tree leader growth is minimal; annual non-native species may be the dominant understory species and will typically be found under the tree canopies. Trace amounts of sagebrush may be present however dead skeletons will be more numerous than living sagebrush. Bunchgrasses may or may not be present. Sandberg bluegrass or mat forming forbs may be present in trace amounts. Bare ground interspaces are large and connected. Soil redistribution is evident.
Pathway a
Community 5.1 to 5.2
Absence of disturbance over time allows for tree cover and density to further increase and out-compete the herbaceous understory species for sunlight and water.
Transition A
State 1 to 2
Trigger: This transition is caused by the introduction of non-native annual weeds, such as cheatgrass, mustard, poverty weed, and halogeton. Slow variables: Over time the annual non-native plants will increase within the community. Threshold: Any amount of introduced non-native species causes an immediate decrease in the resilience of the site. Annual non-native species cannot be easily removed from the system and have the potential to significantly alter disturbance regimes from their historic range of variation.
Transition A
State 2 to 3
Trigger: Inappropriate, long-term grazing of perennial bunchgrasses during growing season would favor shrubs and initiate transition to Community Phase 3.1. Fire would cause a transition to Community Phase 3.2. Slow variables: Long term decrease in deep-rooted perennial grass density. Threshold: Loss of deep-rooted perennial bunchgrasses changes spatial and temporal nutrient cycling and nutrient redistribution, and reduces soil organic matter.
Transition B
State 2 to 4
Trigger: Fire or a failed seeding leads to plant community phase 4.1, inappropriate grazing management that favors shrubs in the presence of non-native annual species leads to community phase 4.2. Slow variables: Increased production and cover of non-native annual species. Threshold: Cheatgrass or other non-native annuals dominate understory.
Transition C
State 2 to 5
Trigger: Time allows for trees to increase. Slow variables: Increased establishment and cover of Utah juniper. Threshold: Trees overtop Wyoming sagebrush and out-compete shrubs for water and sunlight. Shrub skeletons exceed live shrubs with minimal recruitment of new cohorts.
Transition A
State 3 to 4
Trigger: Fire or abusive grazing can eliminate the Sandberg bluegrass understory and transition to community phase 4.1 or 4.2. Slow variable: Increased seed production and cover of annual non-native species. Threshold: Increased, continuous fine fuels modify the fire regime by changing intensity, size and spatial variability of fires. Changes in plant community composition and spatial variability of vegetation due to the loss of perennial bunchgrasses and sagebrush truncate energy capture and impact the nutrient cycling and distribution.
Transition B
State 3 to 5
Trigger: Lack of fire allows for trees to dominate site; may be coupled with inappropriate grazing management that reduces fine fuels. Slow variables: Increased establishment and cover of juniper trees. Threshold: Trees overtop Wyoming sagebrush and out-compete shrubs for water and sunlight. Shrub skeletons exceed live shrubs with minimal recruitment of new cohorts.
Transition A
State 5 to 4
Trigger: Catastrophic crown fire would reduce or eliminate trees to transition the site to 4.1. Tree removal when annual non-natives such as cheatgrass are present would also transition the site to state 4.0. Slow variable: Increased seed production and cover of annual non-native species. Threshold: Increased, continuous fine fuels modify the fire regime by changing intensity, size and spatial variability of fires. Changes in plant community composition and spatial variability of vegetation due to the loss of perennial bunchgrasses and sagebrush truncate energy capture and impact the nutrient cycling and distribution.
Additional community tables
Table 6. Community 1.1 plant community composition
| Group | Common name | Symbol | Scientific name | Annual production (lb/acre) | Foliar cover (%) | |
|---|---|---|---|---|---|---|
|
Grass/Grasslike
|
||||||
| 1 | Primary Perennial Grasses | 204–378 | ||||
| Thurber's needlegrass | ACTH7 | Achnatherum thurberianum | 150–210 | – | ||
| Indian ricegrass | ACHY | Achnatherum hymenoides | 30–90 | – | ||
| needle and thread | HECO26 | Hesperostipa comata | 12–48 | – | ||
| Sandberg bluegrass | POSE | Poa secunda | 12–30 | – | ||
| 2 | Secondary Perennial Grasses | 12–48 | ||||
| blue grama | BOGR2 | Bouteloua gracilis | 3–18 | – | ||
| squirreltail | ELEL5 | Elymus elymoides | 3–18 | – | ||
| western wheatgrass | PASM | Pascopyrum smithii | 3–18 | – | ||
| James' galleta | PLJA | Pleuraphis jamesii | 3–18 | – | ||
| bluebunch wheatgrass | PSSPS | Pseudoroegneria spicata ssp. spicata | 3–18 | – | ||
|
Forb
|
||||||
| 3 | Perennial | 12–90 | ||||
| aster | ASTER | Aster | 5–30 | – | ||
| arrowleaf balsamroot | BASA3 | Balsamorhiza sagittata | 5–30 | – | ||
| tapertip hawksbeard | CRAC2 | Crepis acuminata | 5–30 | – | ||
| fleabane | ERIGE2 | Erigeron | 5–30 | – | ||
| lupine | LUPIN | Lupinus | 5–30 | – | ||
| phlox | PHLOX | Phlox | 5–30 | – | ||
|
Shrub/Vine
|
||||||
| 4 | Primary Shrubs | 132–240 | ||||
| Wyoming big sagebrush | ARTRW8 | Artemisia tridentata ssp. wyomingensis | 120–210 | – | ||
| spiny hopsage | GRSP | Grayia spinosa | 12–30 | – | ||
| 5 | Secondary Shrubs | 27–78 | ||||
| yellow rabbitbrush | CHVI8 | Chrysothamnus viscidiflorus | 3–18 | – | ||
| Nevada jointfir | EPNE | Ephedra nevadensis | 3–18 | – | ||
| antelope bitterbrush | PUTR2 | Purshia tridentata | 3–18 | – | ||
|
Tree
|
||||||
| 6 | Evergreen | 3–12 | ||||
| Utah juniper | JUOS | Juniperus osteosperma | 3–12 | – | ||
Interpretations
Animal community
Livestock Interpretations:
This site is suitable for livestock grazing. Considerations for grazing management including timing, intensity and duration of grazing. Targeted grazing could be used to decrease the density of non-natives. Indian ricegrass is highly palatable to all classes of livestock in both green and cured condition. It supplies a source of green feed before most other native grasses have produced much new growth. Needleandthread provides highly palatable forage, especially in the spring before fruits have developed. Needlegrasses are grazed in the fall only if the fruits are softened by rain. When actively growing, galleta provides good to excellent forage for cattle and horses and fair forage for domestic sheep. Although not preferred, all classes of livestock may use galleta when it is dry. Domestic sheep show greater use in winter than summer months and typically feed upon central portions of galleta tufts, leaving coarser growth around the edges. Galleta may prove somewhat coarse to domestic sheep. Livestock browse Wyoming big sagebrush, but may use it only lightly when palatable herbaceous species are available. Spiny hopsage provides a palatable and nutritious food source for livestock, particularly during late winter through spring. Domestic sheep browse the succulent new growth of spiny hopsage in late winter and early spring. Overgrazing leads to an increase in sagebrush and a decline in understory plants like Thurber's needlegrass, Indian ricegrass and needleandthread grass. Squirreltail or galleta grass will increase temporarily with further degradation. Invasion of annual weedy forbs and cheatgrass could occur with further grazing degradation, leading to a decline in squirreltail and an increase in bare ground. A combination of overgrazing and prolonged drought leads to soil erosion, increased bare ground and a loss in plant production. Wildfire in sites with cheatgrass present could transition to cheatgrass dominated communities. Without management cheatgrass and annual forbs are likely to invade and dominate the site, especially after fire. Although trees are part of the site concept, Utah juniper and/or singleleaf pinyon can increase and eventually dominate this site.
The Thurber’s needlegrass component of this plant community is an important forage source for livestock and wildlife in the arid regions of the West (Ganskopp 1988). Although the seeds are apparently not injurious, grazing animals avoid them when they begin to mature. Sheep, however, have been observed to graze the leaves closely, leaving stems untouched (Eckert and Spencer 1987). Heavy grazing during the growing season has been shown to reduce the basal area of Thurber’s needlegrass (Eckert and Spencer 1987), suggesting that both seasonality and utilization are important factors in management of this plant. A single defoliation, particularly during the boot stage, was found to reduce herbage production and root mass thus potentially lowering the competitive ability of this needlegrass (Ganskopp 1988).
Needleandthread grass is most commonly found on warm/dry soils (Miller et al. 2013). It is not grazing tolerant and will be one of the first grasses to decrease under heavy grazing pressure (Smoliak et al. 1972, Tueller and Blackburn 1974). Heavy grazing is likely to reduce basal area of these plants (Smoliak et al. 1972). With the reduction in competition from deep rooted perennial bunchgrasses, shallower rooted grasses such as Sandberg bluegrass and galleta grass and forbs may increase (Smoliak et al. 1972).
Indian ricegrass is a deep-rooted, cool season perennial bunchgrass that is adapted primarily to sandy soils. Indian ricegrass is a preferred forage species for livestock and wildlife (Cook 1962, Booth et al. 2006). This species is often heavily utilized in winter because it cures well (Booth et al. 2006). It is also readily utilized in early spring, being a source of green feed before most other perennial grasses have produced new growth (Quinones 1981). Booth et al (2006) also notes that the plant does well when utilized in winter and spring. Cook and Child (1971), however, found that repeated heavy grazing reduced crown cover, which may reduce seed production, density, and basal area of these plants. Additionally, heavy early spring grazing reduces plant vigor and stand density (Stubbendieck et al. 1985). In eastern Idaho, productivity of Indian ricegrass was at least 10 times greater in undisturbed plots than in heavily grazed ones (Pearson 1976). Yet, Cook and Child (1971) found significant reduction in plant cover even after 7 years of rest from heavy (90%) and moderate (60%) spring use. The seed crop may be reduced where grazing is heavy (Bich et al. 1995). Tolerance to grazing increases after May, thus spring deferment may be necessary for stand enhancement (Pearson 1964, Cook and Child 1971); however, utilization of less than 60% is recommended. In summary, adaptive management is required to manage this bunchgrass well.
Reduced bunchgrass vigor or density provides an opportunity for galleta and/or Sandberg bluegrass expansion and/or cheatgrass and other invasive species such as halogeton to occupy interspaces. Increased cheatgrass cover leads to increased fire frequency and potentially an annual plant community. Galleta and/or Sandberg bluegrass increases under grazing pressure (Jameson 1962, Tisdale and Hironaka 1981) and is capable of co-existing with cheatgrass. Excessive sheep grazing favors galleta or Sandberg bluegrass; however, where cattle are the dominant grazers, cheatgrass often dominates (Daubenmire 1970). Thus, depending on the season of use, the type of grazing animal, and site conditions, either galleta or Sandberg bluegrass or cheatgrass may become the dominant understory with inappropriate grazing management.
Stocking rates vary over time depending upon season of use, climate variations, site, and previous and current management goals. A safe starting stocking rate is an estimated stocking rate that is fine tuned by the client by adaptive management through the year and from year to year.
Wildlife Interpretations:
A large compliment of wildlife species, including many bird, small and large mammals, and reptile species, depend on or at least partially utilize this ecological site. Large mammals common to this ecological site include mule deer (Odocoileus hemionus) and pronghorn antelope (Antilocarpa americana). Desert bighorn sheep (Ovis canadensis sierra) and Rocky Mountain elk (Cervus elaphus nelsonii canadensis) may occasionally use this ecological site; especially when in search of green forage in the late-winter and early-spring months. Wyoming big sagebrush is preferred browse for wild ungulates. Pronghorn usually browse Wyoming big sagebrush heavily. Spiny hopsage provides a palatable and nutritious food source for big game, particularly during late winter through spring. Thurber’s needlegrass is valuable forage for wildlife. Indian ricegrass is eaten by pronghorn in moderate amounts whenever available. In Nevada, Indian ricegrass is consumed by desert bighorns.
Sagebrush-grassland communities provide critical sage-grouse (Centrocercus urophasianus) breeding and nesting habitats. Sagebrush is a crucial component of their diet year-round, and sage-grouse select sagebrush almost exclusively for cover. Sage-grouse prefer mountain big sagebrush and Wyoming big sagebrush communities to basin big sagebrush communities. In addition to sage-grouse; California quail (Callipepla californica), chukar (Alectoris chukar), mourning dove (Zenaida macroura), Brewers sparrow (Spizella breweri), gray flycatcher (Empidonax wrightii), lark sparrow (Chondestes grammacus), sage sparrow (Amphispiza belli), sage thrasher (Oreoscoptes montanus), and Vesper’s sparrow (Pooecetes gramineus) utilize this ecological site. Indian ricegrass seed provides food for many species of birds. Doves, for example, eat large amounts of shattered Indian ricegrass seed lying on the ground.
This sagebrush-grassland ecological site also provides habitat for small mammals such as: black-tailed (Lepus californicus) and white-tailed jackrabbit (L. townsendii), mountain cottontail (Sylvilagus nuttallii), pygmy rabbit (Brachylagus idahoensis), dark kangaroo mouse (Microdipodops megacephalus), Great Basin ground squirrel (Sermophilus mollis), Great Basin pocket mouse (Perognathus parvus), little pocket mouse (P. longimembris), Merriam’s shrew (Sorex merriami), Northern grasshopper mouse (Onychomys leucogaster), Ord’s kangaroo rat (Dipodomys ordii), sagebrush vole (Lemmiscus curtatus), Western jumping mouse (Zapus princeps), and Wyoming ground squirrel (Urocitellus elegans). A number of heteromyid rodents inhabiting desert rangelands show preference for seed of Indian ricegrass. Indian ricegrass is an important component of jackrabbit diets in spring and summer. In Nevada, Indian ricegrass may dominate jackrabbit diets during the spring through the early summer months. Spiny hopsage is a major food source for black-tailed jackrabbits. The seedlings of spiny hopsage are eaten by a variety of small mammals; including rabbits and mice. This ecological site may also provide a significant insect foraging resource for bats.
A variety of predatory bird, mammal, and reptile species utilize this ecological site. Common mammalian predators to this site are the American badger (Taxidea taxus), bobcat (Lynx rufus), coyote (Canis latrans), kit fox (Vulpes macrotis), cougar or mountain lion (Puma concolor), short-tailed weasel (Mustela ermine) and long-tailed weasel (M. frenata). All of which utilize wildlife species in the area as sources of prey. Predatory reptiles utilizing habitat in this ecological site to catch prey are common sagebrush lizard (Sceloporus graciosus), Great Basin spadefoot (Spea intermontana), Great Basin collared lizard (Crotaphytus bicinctores), long-nosed leopard lizard (Gambelia wislizenii), Western skink (Plestiodon skiltonianus), Great Basin whiptail (Aspidoscelis tigris tigris), Great Basin gopher snake (Pituophis catenifer deserticola), Great Basin rattle snake (Crotalus oreganus lutosus). Avian predators present in the site are the American kestrel (Falco sparverius), common raven (Corvus corax), golden eagle (Aquila chrysaetos), loggerhead shrike (Lanius ludovicianus), Western burrowing owl (Athene cunicularia), Ferruginous Hawk (Buteo regalis), Swainson’s hawk (B. swainsoni), and Prairie falcon (Falco mexicanus).
Changes in soil properties, plant community composition, precipitation, and fire frequency associated with this ecological site could affect the distribution and presence of wildlife species.
Hydrological functions
Runoff is high. Permeability is moderately rapid.
Recreational uses
Aesthetic value is derived from the diverse floral and faunal composition and the colorful flowering of wild flowers and shrubs during the spring and early summer. This site offers rewarding opportunities to photographers and for nature study. This site is used for camping and hiking and has potential for upland and big game hunting.
Other products
Native Americans made tea from big sagebrush leaves. They used the tea as a tonic, an antiseptic, for treating colds, diarrhea, and sore eyes and as a rinse to ward off ticks. Big sagebrush seeds were eaten raw or made into meal. Some Native American peoples traditionally ground parched seeds of spiny hopsage to make pinole flour. Indian ricegrass was traditionally eaten by some Native Americans. The Paiutes used seed as a reserve food source.
Other information
Wyoming big sagebrush is used for stabilizing slopes and gullies and for restoring degraded wildlife habitat, rangelands, mine spoils and other disturbed sites. It is particularly recommended on dry upland sites where other shrubs are difficult to establish. Spiny hopsage has moderate potential for erosion control and low to high potential for long-term revegetation projects. It can improve forage, control wind erosion, and increase soil stability on gentle to moderate slopes. Spiny hopsage is suitable for highway plantings on dry sites in Nevada. Needleandthread grass is useful for stabilizing eroded or degraded sites.
Supporting information
Type locality
| Location 1: White Pine County, NV | |
|---|---|
| Township/Range/Section | T12N R61E S8 |
| General legal description | SE¼NW¼ Section 8, T12N. R61E. MDBM. About 4 miles west of Preston, White Pine County, Nevada. |
Other references
Akinsoji, A. 1988. Postfire vegetation dynamics in a sagebrush steppe in southeastern Idaho, USA. Vegetatio 78:151-155.
Baker, W. L. 2006. Fire and restoration of sagebrush ecosystems. Wildlife Society Bulletin 34:177-185.
Baker, W. L. 2011. Pre-euro-american and recent fire in sagebrush ecosystems. Pages 185-201 in S. T. Knick and J. W. Connelly, editors. Greater sage-grouse: ecology and conservation of a landscape species and its habitats. University of California Press, Berkeley, California.
Balch, J. K., B. A. Bradley, C. M. D'Antonio, and J. Gómez-Dans. 2013. Introduced annual grass increases regional fire activity across the arid western USA (1980–2009). Global Change Biology 19:173-183.
Bich, B.S., J.L. Butler, and C.A. Schmidt. 1995. Effects of differential livestock use of key plant species and rodent populations within selected Oryzopsis hymenoides/Hilaria jamesii communities in Glen Canyon National Recreation Area. The Southwestern Naturalist 40(3):281-287.
Booth, D. T., C. G. Howard, and C. E. Mowry. 2006. 'Nezpar'Indian ricegrass: description, justification for release, and recommendations for use. Rangelands Archives 2:53-54.
Bradley, A. F., N. V. Noste, and W. C. Fischer. 1992. Gen. Tech. Rep. INT-287: Fire ecology of forests and woodlands in Utah. . U.S. Department of Agriculture, Forest Service, Intermountain Research Station, Ogden, UT.
Bunting, S.C., B.M. Kilgore, and C.L. Bushey. 1987. Guidelines for prescribed burning sagebrushgrass rangelands in the northern Great Basin. Gen. Tech. Rep. INT-231. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 33 p.
Caudle, D., J. DiBenedetto, M. Karl, H. Sanchez, and C. Talbot. 2013. Interagency ecological site handbook for rangelands. Available at: http://jornada.nmsu.edu/sites/jornada.nmsu.edu/files/InteragencyEcolSiteHandbook.pdf. Accessed 4 October 2013.
Chambers, J. C., B. A. Roundy, R. R. Blank, S. E. Meyer, and A. Whittaker. 2007. What makes great basin sagebrush ecosystems invasible by Bromus tectorum? Ecological Monographs 77:117-145.
Chambers, J., B. Bradley, C. Brown, C. D’Antonio, M. Germino, J. Grace, S. Hardegree, R. Miller, and D. Pyke. 2013. Resilience to Stress and Disturbance, and Resistance to Bromus tectorum L. Invasion in Cold Desert Shrublands of Western North America. Ecosystems:1-16.
Comstock, J and J. Ehleringer. 1992. Plant adaptation in the Great Basin and Colorado plateau. Western North American Naturalist 52(3):195-215.
Cook, C. W. 1962. An Evaluation of Some Common Factors Affecting Utilization of Desert Range Species. Journal of Range Management 15:333-338.
Cook, C.W. and R.D. Child. 1971. Recovery of desert plants in various states of vigor. Journal of Range Management 24(5):339-343.
Daubenmire, R. 1975. Plant succession on abandoned fields, and fire influences in a steppe area in southeastern Washington. Northwest Science 49:36-48.
Davies, K. W., J. D. Bates, and R. F. Miller. 2006. Vegetation Characteristics across Part of the Wyoming Big Sagebrush Alliance. Rangeland Ecology & Management 59:567-575.
Dobrowolski, J.P., Caldwell, M.M. and Richards, J.H. 1990. Basin hydrology and plant root systems. In: Plant Biology of the Basin and Range. Springer-Verlag Pub., New York, NY.
Fire Effects Information System (Online; http://www.fs.fed.us/database/feis/plants/).
Houghton, J.G., C.M. Sakamoto, and R.O. Gifford. 1975. Nevada’s Weather and Climate, Special Publication 2. Nevada Bureau of Mines and Geology, Mackay School of Mines, University of Nevada, Reno, NV.
Jameson, Donald A. 1962. Effects of burning on a galleta-black grama range invaded by juniper. Ecology. 43(4): 760-763.
Lett, M. S. and A. K. Knapp. 2005. Woody Plant Encroachment and Removal in Mesic Grassland: Production and Composition Responses of Herbaceous Vegetation. The American Midland Naturalist 153:217-231.
Miller, R. F. R. J. T. 2000. The role of fire in juniper and pinyon woodlands: a descriptive analysis. Pages p. 15-30 in Proceedings of the invasive species workshop: the role of fire in the control and spread of invasive species., Tallahassee, Florida.
Miller, R. F., J. C. Chambers, D. A. Pyke, F. B. Pierson, and C. J. Williams. 2013. A review of fire effects on vegetation and soils in the Great Basin Region: response and ecological site characteristics.
National Oceanic and Atmospheric Administration. 2004. The North American Monsoon. Reports to the Nation. National Weather Service, Climate Prediction Center. Available online: http://www.weather.gov/
Noy-Meir, I. 1973. Desert Ecosystems: Environment and Producers. Annual Review of Ecology and Systematics 4:25-51.
Pearson, L.C. 1964. Effect of harvest date on recovery of range grasses and shrubs. Agronomy Journal 56:80-82.
Pearson, L.C. 1976. Primary production in grazed and ungrazed desert communities of eastern Idaho. Ecology 46(3):278-285.
Pierson, F. B., C. J. Williams, P. R. Kormos, S. P. Hardegree, P. E. Clark, and B. M. Rau. 2010. Hydrologic Vulnerability of Sagebrush Steppe Following Pinyon and Juniper Encroachment. Rangeland Ecology & Management 63:614-629.
Quinones, F.A. 1981. Indian ricegrass evaluation and breeding. Bulletin 681. Las Cruces, NM: New Mexico State University, Agricultural Experiment Station. 19 p.
Richards, J.H. and M.M. Caldwell. 1987. Hydralulic lift: Substantial nocturnal water transport between soil layers by Artemisia tridentate roots. Oecologia 73(4):486-489.
Smoliak, S., J. F. Dormaar, and A. Johnston. 1972. Long-Term Grazing Effects on Stipa-Bouteloua Prairie Soils. Journal of Range Management 25:246-250.
Stringham, T.K., P. Novak-Echenique, P. Blackburn, C. Coombs, D. Snyder and A. Wartgow. 2015. Final Report for USDA Ecological Site Description State-and-Transition Models, Major Land Resource Area 28A and 28B Nevada. University of Nevada Reno, Nevada Agricultural Experiment Station Research Report 2015-01. p. 1524.
Stubbendieck, J., J.T. Nichols, and K.K. Roberts. 1985. Nebraska range and pasture grasses (including grass-like plants). E.C. 85-170. Lincoln, NE: University of Nebraska, Department of Agriculture, Cooperative Extension Service. 75 p.
Tisdale, E.W. and M. Hironaka. 1981. The sagebrush-grass region: a review of the ecological literature. Bull. 33. Moscow, ID: University of Idaho, Forest, Wildlife and Range Experiment Station. 31 p.
Tueller, P. T. and W. H. Blackburn. 1974. Condition and Trend of the Big Sagebrush/Needleandthread Habitat Type in Nevada. Journal of Range Management 27:36-40.
USDA-NRCS Plants Database (Online; http://www.plants.usda.gov).
West, N.E. 1994. Effects of fire on salt-desert shrub rangelands. In: Monsen, S.B. and S.G. Kitchen (compilers). Proceedings--ecology and management of annual rangelands; 1992 May 18-22; Boise, ID. Gen. Tech. Rep. INT-GTR-313. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station: Pgs 71-74.
West, N.E. and M.A. Hassan. 1985. Recovery of sagebrush-grass vegetation following wildfire. Journal of Range Management 38(2):131-134.
Wright, H. A. 1971. Why Squirreltail Is More Tolerant to Burning than Needle-and-Thread. Journal of Range Management 24:277-284.
Wright, H. A. 1985. Effects of fire on grasses and forbs in sagebrush-grass communities. Pages 12-21 in Rangeland Fire Effects; A Symposium: Boise, ID, USDI-BLM.
Wright, H. A. and J. O. Klemmedson. 1965. Effect of Fire on Bunchgrasses of the Sagebrush-Grass Region in Southern Idaho. Ecology 46:680-688.
Young, J.A., R.E. Eckert, Jr., R.A. Evans. 1979. Historical perspectives regarding the sagebrush ecosystem. In: The sagebrush ecosystem: a symposium: Proceedings; 1978 April; Logan, UT. Logan, UT: Utah State University, College of Natural Resources: 1-13.
Young, R.P. 1983. Fire as a vegetation management tool in rangelands of the Intermountain Region. In: Monsen, S.B. and N. Shaw (compilers). Managing Intermountain rangelands--improvement of range and wildlife habitats: Proceedings; 1981 September 15-17; Twin Falls, ID; 1982 June 22-24; Elko, NV. Gen. Tech. Rep. INT-157. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: Pgs 18-31.
Contributors
RK
T. Stringham
Rangeland health reference sheet
Interpreting Indicators of Rangeland Health is a qualitative assessment protocol used to determine ecosystem condition based on benchmark characteristics described in the Reference Sheet. A suite of 17 (or more) indicators are typically considered in an assessment. The ecological site(s) representative of an assessment location must be known prior to applying the protocol and must be verified based on soils and climate. Current plant community cannot be used to identify the ecological site.
| Author(s)/participant(s) | P. Novak-Echenique |
|---|---|
| Contact for lead author | State Rangeland Management Specialist |
| Date | 10/28/2013 |
| Approved by | |
| Approval date | |
| Composition (Indicators 10 and 12) based on | Annual Production |
Indicators
-
Number and extent of rills:
Rills are none to rare. A few can be expected on steeper slopes after summer convection storms or rapid snowmelt. These will begin healing during the next growing season. -
Presence of water flow patterns:
Water flow patterns are none to rare. A few can be expected on steeper slopes after summer convectio storms or rapid snowmelt. If present, they are short (<2m), meandering and not connected. -
Number and height of erosional pedestals or terracettes:
Pedestals are none to rare. Occurrence is usually limited to areas of water flow patterns. Frost heaving of shallow rooted plants should not be considered an indicator of soil erosion. -
Bare ground from Ecological Site Description or other studies (rock, litter, lichen, moss, plant canopy are not bare ground):
Bare Ground ± 10-20%, depending on amount of surface rock fragments. -
Number of gullies and erosion associated with gullies:
None -
Extent of wind scoured, blowouts and/or depositional areas:
None -
Amount of litter movement (describe size and distance expected to travel):
Fine litter (foliage from grasses and annual & perennial forbs) expected to move distance of slope length during intense summer convection storms or rapid snowmelt events. Persistent litter (large woody material) will remain in place except during large rainfall events. -
Soil surface (top few mm) resistance to erosion (stability values are averages - most sites will show a range of values):
Soil stability values should be 3 to 6 on most soil textures found on this site. -
Soil surface structure and SOM content (include type of structure and A-horizon color and thickness):
Surface structure is typically moderate, thin platy or fine granular. Soil surface colors are grayish browns and soils are typified by a ochric epipedon. Surface textures are clay loams. Organic matter of the surface 2 to 4 inches is typically less than 1 percent dropping off quickly below. Organic matter content can be more or less depending on micro-topography. -
Effect of community phase composition (relative proportion of different functional groups) and spatial distribution on infiltration and runoff:
Perennial herbaceous plants (especially deep-rooted bunchgrasses [i.e., Thurber's needlegrass & Indian ricegrass]) slow runoff and increase infiltration. Shrub canopy and associated litter break raindrop impact and provide opportunity for snow catch and accumulation on site. -
Presence and thickness of compaction layer (usually none; describe soil profile features which may be mistaken for compaction on this site):
Compacted layers are none. Subangular blocky or massive structure or subsoil argillic horizons are not to be interpreted as compacted. -
Functional/Structural Groups (list in order of descending dominance by above-ground annual-production or live foliar cover using symbols: >>, >, = to indicate much greater than, greater than, and equal to):
Dominant:
Reference State: Deep-rooted, cool season, perennial bunchgrasses (Indian ricegrass & Thurber's needlegrass)Sub-dominant:
Tall shrubs (big sagebrush & spiny hopsage) >> associated shrubs >shallow-rooted, cool season, perennial bunchgrasses = fibrous, shallow-rooted, cool season, perennial forbs = deep-rooted, cool season, perennial forbs > warm season rhizomatous grasses.Other:
Cool season, rhizomatous grasses, annual forbs, and evergreen trees.Additional:
With an extended fire return interval, the shrub and tree component will increase at the expense of the herbaceous component. Singleleaf pinyon and Utah juniper may eventually dominate the site and the understory component will be greatly reduced. -
Amount of plant mortality and decadence (include which functional groups are expected to show mortality or decadence):
Dead branches within individual shrubs common and standing dead shrub canopy material may be as much as 20% of total woody canopy; some of the mature bunchgrasses (<10%) have dead centers. -
Average percent litter cover (%) and depth ( in):
Between plant interspaces (±10-15%) and litter depth is <¼ inch. -
Expected annual annual-production (this is TOTAL above-ground annual-production, not just forage annual-production):
For normal or average growing season (through June) ±600 lbs/ac. Favorable years + 800 lbs/ac and unfavorable years + 350 lbs/ac. -
Potential invasive (including noxious) species (native and non-native). List species which BOTH characterize degraded states and have the potential to become a dominant or co-dominant species on the ecological site if their future establishment and growth is not actively controlled by management interventions. Species that become dominant for only one to several years (e.g., short-term response to drought or wildfire) are not invasive plants. Note that unlike other indicators, we are describing what is NOT expected in the reference state for the ecological site:
Potential invaders include cheatgrass, Russian thistle, annual mustards and singleleaf pinyon. With an extended fire return interval, Utah juniper and singleleaf pinyon will increase on this site eventually dominating it. -
Perennial plant reproductive capability:
All functional groups should reproduce in average (or normal) and above average growing season years. Little growth or reproduction occurs during extreme or extended drought periods.
Print Options
Sections
Font
Other
The Ecosystem Dynamics Interpretive Tool is an information system framework developed by the USDA-ARS Jornada Experimental Range, USDA Natural Resources Conservation Service, and New Mexico State University.
Click on box and path labels to scroll to the respective text.