
Natural Resources
Conservation Service
Ecological site F028BY067NV
POTR5/SYOR2/BRMA4-ELTR7
Accessed: 04/15/2026
General information
Provisional. A provisional ecological site description has undergone quality control and quality assurance review. It contains a working state and transition model and enough information to identify the ecological site.
Figure 1. Mapped extent
Areas shown in blue indicate the maximum mapped extent of this ecological site. Other ecological sites likely occur within the highlighted areas. It is also possible for this ecological site to occur outside of highlighted areas if detailed soil survey has not been completed or recently updated.
MLRA notes
Major Land Resource Area (MLRA): 028B–Central Nevada Basin and Range
MLRA 28B occurs entirely in Nevada and comprises about 23,555 square miles (61,035 square kilometers). More than nine-tenths of this MLRA is federally owned. This area is in the Great Basin Section of the Basin and Range Province of the Intermontane Plateaus. It is an area of nearly level, aggraded desert basins and valleys between a series of mountain ranges trending north to south. The basins are bordered by long, gently sloping to strongly sloping alluvial fans. The mountains are uplifted fault blocks with steep sideslopes. Many of the valleys are closed basins containing sinks or playas. Elevation ranges from 4,900 to 6,550 feet (1,495 to 1,995 meters) in the valleys and basins and from 6,550 to 11,900 feet (1,995 to 3,630 meters) in the mountains.
The mountains in the southern half are dominated by andesite and basalt rocks that were formed in the Miocene and Oligocene. Paleozoic and older carbonate rocks are prominent in the mountains to the north. Scattered outcrops of older Tertiary intrusives and very young tuffaceous sediments are throughout this area. The valleys consist mostly of alluvial fill, but lake deposits are at the lowest elevations in the closed basins. The alluvial valley fill consists of cobbles, gravel, and coarse sand near the mountains in the apex of the alluvial fans. Sands, silts, and clays are on the distal ends of the fans.
The average annual precipitation ranges from 4 to 12 inches (100 to 305 millimeters) in most areas on the valley floors. Average annual precipitation in the mountains ranges from 8 to 36 inches (205 to 915 millimeters) depending on elevation. The driest period is from midsummer to midautumn. The average annual temperature is 34 to 52 degrees F (1 to 11 degrees C). The freeze-free period averages 125 days and ranges from 80 to 170 days, decreasing in length with elevation.
The dominant soil orders in this MLRA are Aridisols, Entisols, and Mollisols. The soils in the area dominantly have a mesic soil temperature regime, an aridic or xeric soil moisture regime, and mixed or carbonatic mineralogy. They generally are well drained, loamy or loamyskeletal, and shallow to very deep.
Nevada’s climate is predominantly arid, with large daily ranges of temperature, infrequent severe storms and heavy snowfall in the higher mountains. Three basic geographical factors largely influence Nevada’s climate: continentality, latitude, and elevation. The strong continental effect is expressed in the form of both dryness and large temperature variations. Nevada lies on the eastern, lee side of the Sierra Nevada Range, a massive mountain barrier that markedly influences the climate of the State. The prevailing winds are from the west, and as the warm moist air from the Pacific Ocean ascend the western slopes of the Sierra Range, the air cools, condensation occurs and most of the moisture falls as precipitation. As the air descends the eastern slope, it is warmed by compression, and very little precipitation occurs. The effects of this mountain barrier are felt not only in the West but throughout the state, as a result the lowlands of Nevada are largely desert or steppes.
The temperature regime is also affected by the blocking of the inland-moving maritime air. Nevada sheltered from maritime winds, has a continental climate with well-developed seasons and the terrain responds quickly to changes in solar heating. Nevada lies within the midlatitude belt of prevailing westerly winds which occur most of the year. These winds bring frequent changes in weather during the late fall, winter and spring months, when most of the precipitation occurs.
To the south of the mid-latitude westerlies, lies a zone of high pressure in subtropical latitudes, with a center over the Pacific Ocean. In the summer, this high-pressure belt shifts northward over the latitudes of Nevada, blocking storms from the ocean. The resulting weather is mostly clear and dry during the summer and early fall, with occasional thundershowers. The eastern portion of the state receives noteworthy summer thunderstorms generated from monsoonal moisture pushed up from the Gulf of California, known as the North American monsoon. The monsoon system peaks in August and by October the monsoon high over the Western U.S. begins to weaken and the precipitation retreats southward towards the tropics (NOAA 2004).
Ecological site concept
This forest site occurs on mountain sideslopes. Slope gradients of 15 to 50 percent are typical. Elevations are 7000 to about 10,800 feet.
The soils associated with this site are very deep, well drained and formed in colluvium and residuum derived from quartzite. Soils are characterized by a mollic epipedon from soil surface to 32cm and greater than 35% rock fragments by volume distributed throughout the profile.
The reference state is composed of one to several quaking aspen clones. An overstory canopy of about 20 to 35 percent is assumed to be representative. Understory composition is about 40% grass, 30% forbs and 30% shrubs. Mountain brome, wheatgrasses, meadowrue, snowberry and currant are common in the understory and production ranges from 400 to 800 pounds per acre. Overstory tree canopy is 100 percent quaking aspen.
Important abiotic factors associated with this site include run –on moisture from the surrounding landscape and the cool soil temperatures. Run-on moisture results in increased annual production and therefore increased organic matter that develops the thicker mollic epipdon consistent with aspen- dominated ecological sites.
Similar sites
| F028BY055NV |
POTR5-ABCO/LEKI2-BRMA4-ELTR7 This site occurs on steep mountain sideslopes. Aspen site index is higher. Understory dominated by spike fescue, bluebunch wheatgrass, mountain brome, and slender wheatgrass. White fir occurs in the reference state. |
|---|---|
| F028BY025NV |
Mountain Stream Terrace This site occurs on nearly level to gently sloping mountain valleys and along mountain stream terraces. Understory is dominated by willow, Wood's rose, slender wheatgrass, Nevada bluegrass and tufted hairgrass. Site index and understory production are higher. |
Table 1. Dominant plant species
| Tree |
(1) Populus tremuloides |
|---|---|
| Shrub |
(1) Symphoricarpos oreophilus |
| Herbaceous |
(1) Bromus marginatus |
Physiographic features
This site typically occurs on backslopes and shoulder positions of mountains. Slopes range from 8 to 75 percent, but gradients of 15 to 50 percent are typical. Elevations are 7000 to about 10,800 feet.
Table 2. Representative physiographic features
| Landforms |
(1)
Mountain
|
|---|---|
| Elevation | 7,000 – 10,800 ft |
| Slope | 15 – 50% |
| Aspect | Aspect is not a significant factor |
Climatic features
Nevada’s climate is semi-arid. In general it is characterized by cold, moist winters and warm, dry summers.
Average annual precipitation ranges from 16 to over 20 inches. Mean annual air temperature is about 38 to 42 degrees F. The average growing season is 30 to 50 days. Weather stations with a long term data record are currently not available for this ecological site. Associated climate data will be updated when information becomes available.
Table 3. Representative climatic features
| Frost-free period (average) | 40 days |
|---|---|
| Freeze-free period (average) | 0 days |
| Precipitation total (average) | 16 in |
Figure 2. Annual precipitation pattern
Figure 3. Annual average temperature pattern
Influencing water features
Influencing water features are not associated with this site.
Soil features
The soils associated with site are very deep, well drained and formed in residuum/colluvium derived from quartzite. The soil profile is characterized by an organic horizon 5cm thick over a mollic epipedon from 5cm to 32cm and greater than 35% rock fragments by volume. The soil surface is covered with a layer of partially decomposed organic material consisting of aspen leaves, twigs and grass residues. Soil temperature regime is cryic and soil moisture regime is xeric. The potential for sheet and rill erosion is slight to moderate depending on slope. The soil series associated with this site is Berrycreek.
Berrycreek is a loamy-skeletal, mixed, superactive Xeric Haplocryolls. Diagnostic horizons include a mollic epipedon from 5 to 32cm, and a cambic horizon from 32 to 152cm. Clay content in the particle size control section average 18 to 27 percent. Rock fragments range from 45 to 65 percent, consisting mainly of gravel. Reaction is neutral or slightly acid. Soils are non-effervescence and are derived from quartzite.
Table 4. Representative soil features
| Parent material |
(1)
Colluvium
–
quartzite
|
|---|---|
| Surface texture |
(1) Very gravelly loam (2) Gravelly silt loam (3) Very gravelly |
| Family particle size |
(1) Loamy |
| Drainage class | Well drained |
| Permeability class | Slow to moderate |
| Soil depth | 55 – 80 in |
| Surface fragment cover <=3" | 9 – 35% |
| Surface fragment cover >3" | Not specified |
| Available water capacity (0-40in) |
4 – 6 in |
| Calcium carbonate equivalent (0-40in) |
Not specified |
| Electrical conductivity (0-40in) |
2 mmhos/cm |
| Sodium adsorption ratio (0-40in) |
Not specified |
| Soil reaction (1:1 water) (0-40in) |
6.1 – 7.3 |
| Subsurface fragment volume <=3" (Depth not specified) |
40 – 65% |
| Subsurface fragment volume >3" (Depth not specified) |
5 – 15% |
Ecological dynamics
An ecological site is the product of all the environmental factors responsible for its development and it has a set of key characteristics that influence a site’s resilience to disturbance and resistance to invasives. Key characteristics include 1) climate (precipitation, temperature), 2) topography (aspect, slope, elevation, and landform), 3) hydrology (infiltration, runoff), 4) soils (depth, texture, structure, organic matter), 5) plant communities (functional groups, productivity), and 6) natural disturbance regime (fire, herbivory, etc.) (Caudle et al. 2013). Biotic factors that influence resilience include site productivity, species composition and structure, and population regulation and regeneration (Chambers et al. 2013).
Common disturbances in aspen stands include fire, insect and disease outbreaks, wind storms and avalanches. Mature aspen stands (80 to 100 years) can reach heights up to 100 feet depending on the site. Most stands contain a variety of medium-high shrubs and tall herbs in the understory (DeByle and Winokur 1985). Historically wildfire maintained the dynamics of these communities, but with fire suppression mature aspen stands can be susceptible to stand decline. Typically as stands begin to decline canopy openings allow aspen suckers and saplings to regenerate the stand. As aspen trees mature and tree canopy begins to close the perennial understory becomes dominated by shade tolerant species. Pathogen/insect incidence can build-up in aspen stands causing their decadence. Pathogens and insects can decimate decadent aspen allowing infill of conifers.
Aspen sustainability is threatened by limited aspen regeneration due to the lack of disturbance, shading by conifer trees and/or herbivory. Conifers, when present, are shade tolerant and can eventually increase and overtop the aspen trees. The increase in conifers can be attributed to both fire suppression and grazing pressure by livestock and wildlife (Potter 2005, Strand et al. 2009, Bartos and Campbell 1998). Fossorial animals also create or enhance germination sites for conifer seeds (Whitesides 2012). Aspen and their root systems persist through conifer infill, but diminish as conifers predominate. Overstory clearing, whether in small gaps or in large openings, provides the needed light for aspen suckers to sprout (Shepperd et al 2006). A limited aspen root system resulting from previous conifer dominance and/or persistent shading from surrounding uncut trees may require additional disturbance to initiate suckering. Additional management actions such as root ripping may be needed to stimulate root suckering (Shepperd et al 2006). Continuous browsing by livestock or wildlife may also limit aspen regeneration. Herbivory can reduce community resilience and alter future aspen cover (Rogers et al 2013).
There are many environmental factors that can contribute to stand decline or die-off. The major underlying cause can be attributed to tree and/or stand stress. Aspen stands have also shown some sensitivity to drought (Hogg et al 2008). Drought stress and tree-to-tree competition exacerbate insect damage and changes in community phases. Drought, low soil oxygen, and cold soil temperatures all limit soil water uptake and can contribute to xylem cavitation. Cavitation causes much of the aspen die-off but the created stress can also leave the stand open to secondary factors such as wood boring insects and fungal pathogens (Frey et al. 2004). Drought has been attributed to the decline and death of aspen trees, but also contributes to secondary factors such as insects (Frey et al. 2004).
Using a habitat model Strand et al. 2009 computed aspen occurrence probability across the landscape of the Owyhee Plateau. They visited 41 sites where they modeled aspen occurrence; 37% they found dead aspen stems with no aspen regeneration, 51% had scattered aspen and aspen was regenerating in forest gaps, and 12% there was no evidence that aspen had ever occurred on or near the site. Their aspen successional model theorized that non-producing aspen stands can be permanently converted to a conifer stand and the aspen clone can be lost. They estimated that over 60% of aspen woodlands have been or are in the process of converting to conifer woodlands within 80to 200 years. Whether or not these stands can be converted back to aspen with disturbance is inconclusive.
Aspen stands possess three characteristics that provide suitable sites for invasive plants: 1) deep, rich soils, 2) proximity to moist meadows and riparian areas with open water, 3) their dependency on disturbance and open light. This site has moderate resilience to disturbance and resistance to invasion. Human disturbance associated with recreation and animal (domestic and wildlife) disturbance may lead to the spread of invasive species such as Kentucky bluegrass (Poa pratensis), common dandelion (Taraxacum officinale) and thistles (Cirsium sp.). Additionally, this ecological site is moderately resilient and resistant due to productive soils, additional soil moisture and aspens ability to sprout following fire or other stand or tree removal processes. Three stable states have been identified for this site, a reference state, current potential state and a third state where conifers have encroached and dominated the site. The research is inconclusive if these conifer dominated aspen stands can regenerate with fire.
Fire Ecology:
Wildfire is recognized as a natural disturbance that influenced the structure and composition of the vegetation of the Reference State. Specific fire intervals are dependent upon surrounding vegetation communities. Intense fires that kill the aspen overstory usually stimulate abundant suckering (DeByle and Winokur 1985). Although aspen stands rely on fire for successful regeneration, aspen stands don’t readily carry fire (Fechner and Barrows 1976, Debyle and Winokur 1985, Debyle et al. 1987). It is hypothesized that many of the fires that maintained these communities were set by the Native population, who used fire to manage plant communities for human benefit (Kay 1997). The tree itself is extremely fire sensitive (Baker 1925); with its thin bark most aspens are top-killed by fire, and those left with scarring are usually killed within the next growing season from rot and disease (Bradley et al. 1992,Davidson et al. 1959, Meinecke 1929). Periodic wildfires prevent over-mature aspen stands and maintain a naturally stratified mosaic of even-aged aspen communities in various stages of successional development. Uneven-aged stands form under stable conditions where the overstory gradually disintegrates with disease or age, and is replaced by aspen suckers. Historic heavy grazing has been attributed to the reduction of fine fuels within stands; without the fuels to burn fires seldom occur within aspen forests (DeByle and Winokur 1985). Fire suppression or exclusion will result in loss of residual aspen and decreased potential for aspen sprouts. Exclusion will eventually result in older classes of insect-susceptible conifers thereby increasing the risk of catastrophic fires because of fuel-loading and ladder fuels.
Snowberry has been noted to regenerate well and exceed pre-burn biomass in the third season after fire (Merrill et al. 1982). Wax currant, a minor component of this site, is known as a weak sprouter from the root crown but usually regenerates from soil stored seeds after fire. It is susceptible to fire kill and rarely survives fire (Crane and Fischer 1986). If balsamroot or mules ear is common before fire, these plants will increase after fire or with heavy grazing (Wright 1985).
The effect of fire on bunchgrasses relates to culm density, culm-leaf morphology, and the size of the plant. The initial condition of bunchgrasses within the site along with seasonality and intensity of the fire all factor into the individual species response. For most forbs and grasses the growing points are located at or below the soil surface providing relative protection from disturbances which decrease above ground biomass, such as grazing or fire. Thus, fire mortality is more correlated to duration and intensity of heat which is related to culm density, culm-leaf morphology, size of plant and abundance of old growth (Wright 1971, Young 1983).
Mountain brome the dominant grass on this site is a robust, coarse-stemmed, short lived perennial bunchgrass that can grow from 1 to 5 feet in height (Dayton 1937, Tilley et al. 2004). Mountain brome significantly decreases after burning (Nimir and Payne 1978). Slender wheatgrass, a sub-dominant grass on this site, may increase after fire. In a study by Nimir and Payne (1978) slender wheatgrass increased significantly in burned than in non-burned sites, although the species did not appear in measurable quantities until mid-July.
Slender wheatgrass has moderately to densely clustered leaves and stems. Moderate severity fires will top kill plants and some tillers. Fire damage and plant response to fire is dependent on growth form and fire severity. Spring fires may cause more damage, while moderate severity summer and fall fires do not usually effect slender wheatgrass. Post fire recovery of this species is rapid (Howard 1992). Sedges are a common in the herbaceous understory. Generally sedges have rhizomes and only top-killed by wildfire. Muttongrass (Poa fendleriana), also a minor component on this site, is top killed by fire but will resprout after low to moderate severity fires. A study by Vose and White (1991) in an open sawtimber site, found minimal difference in overall effect of burning on muttongrass.
State and transition model
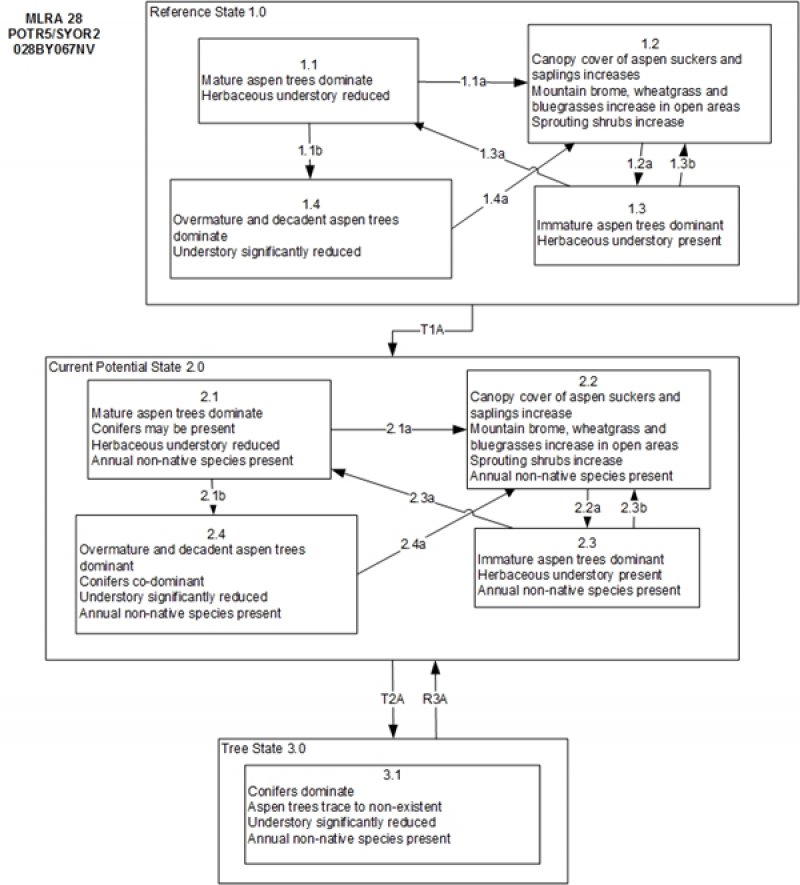
Figure 4. State and Transition Model

Figure 5. Legend
More interactive model formats are also available.
View Interactive Models
More interactive model formats are also available.
View Interactive Models
Click on state and transition labels to scroll to the respective text
Ecosystem states
State 1 submodel, plant communities
State 2 submodel, plant communities
State 3 submodel, plant communities
State 1
Reference State
The Reference State 1.0 is a representative of the natural range of variability under pristine conditions. This site has four general community phases; a mature woodland phase, a sucker/sapling phase, an immature woodland phase and an over mature woodland phase. State dynamics are maintained by interactions between climatic patterns and disturbance regimes. Negative feedbacks enhance ecosystem resilience and contribute to the stability of the state. These include the presence of all structural and functional groups, fuel loads, and retention of organic matter and nutrients. Plant community phase changes are primarily driven by fire, periodic drought and/or insect or disease attack. Periodic wildfires prevent over-mature aspen stands and maintain a naturally stratified mosaic of even-aged aspen stands in various stages of successional development. Many aspen stands are even-aged because of the rapid reproduction by suckering after a major disturbance such as fire. In the absence of fire, harvest, or similar events, aspen stands become uneven-aged. Uneven-aged stands form under stable conditions where the overstory gradually disintegrates with disease or age, and is replaced by suckers. Uneven-aged stands also occur where individual clones gradually expand into adjacent grasslands or shrublands. Harvesting/cutting of older age classes (that are more suspectible to insects) may minimize damage and larger clear-cut areas may retard conifer infill.
Community 1.1
Community Phase
This site is composed of one to several quaking aspen clones, each with a common genetic makeup and individual phenological and physiological characteristics. An overstory canopy of 20 to about 35 percent is assumed to be representative of tree dominance on this site in the pristine environment. These aspen communities are multi-layered. Medium height shrubs and tall herbs frequently form a rather open and intermittent layer beneath the tree canopy. An even lower herbaceous layer is always part of the understory. Sufficient light is able to penetrate the canopies to support abundant understory vegetation. Overstory tree canopy is 100 percent quaking aspen. Mountain brome, slender wheatgrass, streambank wheatgrass, meadowrue, snowberry, sedges and currant are common understory species associated with this site. Understory vegetative composition is about 55 percent grasses, 15 percent forbs and 30 percent shrubs and young trees when the average overstory canopy is medium (20 to 35 percent). Average understory production ranges from 400 to 800 pounds per acre with a medium canopy cover. Understory production includes the total annual production of all species within 5m of the ground surface.
Forest overstory. The visual aspect and vegetal structure are dominated by single-storied aspen that have reached or are near maximal heights for the site. Tree heights range from 30 to 80 feet, depending upon site and clonal genotype. Tree canopy cover ranges from 20 to 35 percent. Trees have developed tall, straight, clear stems with short, narrow dome-like crowns that develop greater at the edge of the stand. Despite considerable forage production in most aspen communities, the overstory trees do compete with the undergrowth plants for moisture, light, nutrients and space. Consequently, adjacent vegetation types lacking such overstory competition will normally produce more forage than the understory of the aspen forest. Vegetative shoots and/or saplings of aspen occur in the understory, but they are inconspicuous and have a high mortality rate.
Forest understory. Understory vegetative composition is about 50 percent grasses, 20 percent forbs and 30 percent shrubs and young trees when the average overstory canopy is medium (20 to 35 percent). Average understory production ranges from 400 to 800 pounds per acre with a medium canopy cover. Understory production includes the total annual production of all species within 5m of the ground surface.
Figure 6. Annual production by plant type (representative values) or group (midpoint values)
Table 5. Annual production by plant type
| Plant type | Low (lb/acre) |
Representative value (lb/acre) |
High (lb/acre) |
|---|---|---|---|
| Grass/Grasslike | 220 | 330 | 440 |
| Shrub/Vine | 92 | 138 | 184 |
| Forb | 60 | 90 | 120 |
| Tree | 28 | 42 | 56 |
| Total | 400 | 600 | 800 |
Community 1.2
Community Phase
Vegetation consists of grasses, forbs and a few shrubs in association with aspen suckers. Quaking aspen suckers are evident. If the aspen stand is healthy, this stage will only last from one to two years. However, if competing brush and herbaceous plants grow for a full season before aspen suckers emerge, or with excessive herbivory from large ungulates such as elk, a reduction in growth and survival of aspen suckers may occur. Early growth of quaking aspen suckers ranges from less than 1 foot to more than 3 feet per year for shoots having good competitive position. In the absence of disturbance, suckers develop into saplings.
Forest understory. Herbaceous vegetation and woody shrubs, including aspen suckers, dominate the site. The understory vegetation consists of grasses, forbs and shrubs in association with tree saplings. Understory production ranges from 600 to 1300 pounds per acre.
Community 1.3
Community Phase
Rapid growth of the aspen trees, both in height and canopy cover quickly results in an immature aspen forest. Aspen stands are self-thinning, especially at young ages. After the canopy closes, trees stratify into crown classes quickly, despite genetic uniformity within clones. Understory vegetation is moderately influenced by a tree overstory canopy. Growth of the aspen begins to slow and there is a fairly continual adjustment of trees to growing space. As competition becomes intense enough to affect the diameter growth of dominants, mortality quickly reduces the number of trees in the lower crown classes.
Forest overstory. The visual aspect and vegetal structure are dominated by aspen ranging from about 10 to 20 feet in height, and having a diameter at breast height of about 2 to 4 inches. Understory vegetation is moderately influenced by a tree overstory canopy up to 20 percent.
Forest understory. Understory production and composition are influenced by the overstory. Shade tolerant forbs and a few grasses increase in the understory, and the grasses and shrubs that dominate in the adjacent rangeland will occur in lesser amounts.
Community 1.4
Community Phase
In the absence of wildfire or other naturally occurring disturbances, the tree canopy on this site can become very dense. This stage is normally dominated by aspen that have reached maximal height. Aspen trees may be decadent. In the absence of disturbance, over-mature, even-aged aspen stands slowly die. The canopy opens up, and otherwise inconspicuous aspen suckers survive and grow in the openings. Tree canopy cover is greater than 35 percent. Understory production is strongly influenced by the overstory, as is species composition. Shade tolerant forbs and a few grasses dominate the understory. Aspen may be represented weakly in the understory.
Forest overstory. Trees have straight, clear stems with short, high-rounded crowns. Tree canopy cover is greater than 35 percent.
Forest understory. Understory vegetation is greatly influenced by a tree overstory canopy greater than 40 percent. Shade tolerant species dominate the understory.
Pathway a
Community 1.1 to 1.2
Fire would reduce the mature aspen and allow for the suckers, saplings and the herbaceous understory to increase.
Pathway b
Community 1.1 to 1.4
Time and lack of disturbance allows the canopy of aspen trees to close reducing regeneration. With time conifer trees in the understory are able to mature further increasing competition with aspen.
Pathway a
Community 1.2 to 1.3
Time and lack of disturbance, release from herbivory will allow for the aspen suckers to mature
Pathway a
Community 1.3 to 1.1
Time and lack of disturbance allows the tallest aspen trees to mature.
Pathway b
Community 1.3 to 1.2
Fire, insects, disease or wind damage can reduce the aspen canopy allowing herbaceous community and aspen suckers to increase.
Pathway a
Community 1.4 to 1.2
Fire removes or drastically reduces tree canopy allowing for aspen suckers to increase. Aspen suckering is stimulated in openings created by tree mortality.
State 2
Current Potential State
This state is similar to the Reference State 1.0 with four similar community phases. Ecological function has not changed, however the resiliency of the state has been reduced by the presence of invasive weeds. Non-natives may increase in abundance but will not become dominant within this State. These non-natives can be highly flammable and can promote fire where historically fire had been infrequent. Negative feedbacks enhance ecosystem resilience and contribute to the stability of the state. These feedbacks include the presence of all structural and functional groups, low fine fuel loads, and retention of organic matter and nutrients. Positive feedbacks decrease ecosystem resilience and stability of the state. These include the non-natives’ high seed output, persistent seed bank, rapid growth rate, ability to cross pollinate, and adaptations for seed dispersal. Harvesting/cutting of older age classes (that are more susceptible to insects) may minimize damage and larger clear-cut areas may retard conifer infill. Season-long herbivory will diminish/eliminate successful aspen establishment and may accelerate establishment of conifers and other woody species. Aspen stands will generally establish is domestic and wild herbivory is restricted to use of leaders on less than 40% of the number of suckers present.
Community 2.1
Community Phase

Figure 7. POTR5 (F028BY067NV) Phase 2.1 P.Novak-Echenique July 2013

Figure 8. POTR5 (F028BY067NV) T. Stringham July 2012
This community phase is similar to the Reference State Community Phase 1.1, with the presence of non-native species in trace amounts such as common dandelion, thistle and cheatgrass. Conifers of different age classes may be present. The visual aspect and vegetal structure are dominated by multi-layered aspen. Mature tree heights range from 30 to 80 feet, depending upon site. Tree canopy cover ranges from 20 to about 35 percent.
Forest overstory. The visual aspect and vegetal structure are dominated by single-storied aspen that have reached or are near maximal heights for the site. Tree heights range from 30 to 80 feet, depending upon site and clonal genotype. Tree canopy cover ranges from 20 to 35 percent. Trees have developed tall, straight, clear stems with short, narrow dome-like crowns that develop greater at the edge of the stand.
Forest understory. Vegetative shoots and/or saplings of aspen occur in the understory, but they are inconspicuous and have a high mortality rate. Understory vegetation is severely reduced due to competition for moisture, light, nutrients and space. Understory production ranges from 400 to 800 pounds per acre.
Community 2.2
Community Phase
Vegetation consists of grasses, forbs and a few shrubs in association with tree saplings. Quaking aspen suckers are evident. If the aspen stand is healthy, these first two stages will only last from one to two years. However, if competing brush and herbaceous plants grow for a full season before aspen suckers emerge sucker survival and growth may be reduced. With excessive grazing from large ungulates such as elk and cattle, a reduction in growth and survival of aspen suckers may occur, this may last until season of grazing is changed, or grazing is reduced/excluded. Early growth of quaking aspen suckers ranges from less than 1 foot to more than 3 feet per year for shoots having good competitive position. Non-native species are stable to increasing within the community.
Forest understory. Vegetation consists of grasses, forbs, sprouting shrubs and aspen suckers Understory production ranges from 600 to 1300 pounds per acre.
Community 2.3
Community Phase

Figure 9. POTR5 (F028BY067NV) T. Stringham July 2013
The visual aspect and vegetal structure are dominated by aspen ranging from about 10 to 20 feet in height, and having a diameter at breast height of about 2 to 4 inches. Understory vegetation is moderately influenced by a tree overstory canopy of <20 percent. Growth of the aspen begins to slow and there is a fairly continual adjustment of trees to growing space. As competition becomes intense enough to affect the diameter growth of dominants, mortality quickly reduces the number of trees in the lower crown classes.
Forest overstory. The visual aspect and vegetal structure are dominated by aspen trees 10-20 feet in height. Canopy cover is <20 percent and comprised entirely of aspen.
Forest understory. Understory vegetation is dominated by grasses, forbs, shrubs and aspen suckers. Understory is moderately influenced by a tree overstory canopy.
Community 2.4
Community Phase
In the absence of wildfire or other naturally occurring disturbances tree canopy on this site can become very dense. This stage is normally dominated by aspen that have reached maximal heights and ages. Aspen trees have straight, clear stems with short, high-rounded crowns. In the absence of disturbance, over-mature, even-aged aspen stands slowly die. Conifers may comprise as much as 50 percent of the total tree canopy in stable, over-mature, aspen stands. Tree canopy cover is commonly more than 40 percent. Understory production is strongly influenced by the overstory, as is species composition. Shade tolerant forbs and a few grasses dominate the understory.
Forest overstory. Tree canopy cover is greater than 40 percent. Aspen trees have straight, clear stems with short, high-rounded crowns. Conifers may comprise as much as 50 percent of the total tree canopy.
Forest understory. Understory production and composition are strongly influenced by the overstory competition for light, moisture and nutrients. Shade tolerant forbs and a few grasses dominate the understory.
Pathway a
Community 2.1 to 2.2
Fire would reduce the mature aspen and allow for the suckers, saplings and the herbaceous understory to increase. Annual non-natives are likely to increase after fire.
Pathway b
Community 2.1 to 2.4
Time and lack of disturbance reduces aspen regeneration and will allow for conifers in the understory to mature and dominate the site.
Pathway a
Community 2.2 to 2.3
Time and lack of disturbance, changing of grazing season or grazing reduction/exclusion will allow for the aspen suckers to mature
Pathway a
Community 2.3 to 2.1
Time and lack of disturbance allows the tallest aspen trees to mature.
Pathway b
Community 2.3 to 2.2
Fire, insects, disease or wind damage can reduce the aspen canopy allowing the understory herbaceous community and aspen suckers to increase.
Pathway a
Community 2.4 to 2.2
Fire, or equivalent such as clearcutting/harvesting of the conifers would allow for the aspen suckers to increase.
State 3
Tree State
This state is characterized by one community phase dominated by Rocky Mountain white fir. Limber pine may also be present. Aspen may be present in trace amounts however trees are decadent and little to no regeneration is occurring. The conifer stand may be a variety of ages depending on harvesting and cutting history and natural regeneration. Aspen and potential for root suckering has been eliminated due to repeated stress from the management regime. Duff build-up can retard conifer germination while fossorial animal activity can provide conifer germination sites. Occurrence of surface fire scorches new seedlings but usually does not kill older conifer trees with thicker bark. Understory vegetation is sparse. Negative feedbacks enhance ecosystem resilience and contribute to the stability of the state. These feedbacks include the dense canopy cover of conifer creating a shade rich environment that facilitates the germination and establishment of conifers and retards the growth and suckering of aspen. Positive feedbacks decrease ecosystem resilience and stability of the state. These include high fuel loads from canopy closure and dead and down wood leading to the potential for stand-replacing fire.
Community 3.1
Community Phase
This community phase is dominated by Rocky Mountain white fir. Limber pine may also be present. Aspen trees may be present but show decadence and are significantly reduced. Understory vegetation is reduced due to competition of the overstory canopy. Annual non-native species may be present.
Forest overstory. In the absence of wildfire or other naturally occurring disturbances, the tree canopy on this site can become dense. Tree canopy may exceed 40 percent and more than half is comprise of white fire or other conifers.
Forest understory. Understory production and composition are strongly influenced by the overstory. Shade tolerant forbs and a few grasses dominate the understory.
Transition A
State 1 to 2
Trigger: This transition is caused by the introduction of non-native plants, such as Kentucky bluegrass, smooth brome, thistles and common dandelion. Conifers may also invade. Slow variables: Over time the annual non-native species and conifers will increase within the community. Threshold: Any amount of introduced non-native species causes an immediate decrease in the resilience of the site. A change in the natural fire return interval allows conifers to increase.
Transition A
State 2 to 3
Trigger: Time and a lack of disturbance allow conifer trees to establish, grow and mature. Slow variables: Over time the abundance and size of trees will increase. Threshold: Conifer composition is greater than 60% of the stand and conifer height exceeds aspen height. Aspen are decadent and dying with little to no regeneration. Little understory vegetation remains due to competition with trees for site resources.
Restoration pathway A
State 3 to 2
Prescribed fire or mechanical removal of trees potentially coupled with root ripping to stimulate suckering. Planting of aspen stock may also be required. Restoration of an aspen-dominated site may be extremely difficult and not assured.
Additional community tables
Table 6. Community 1.1 plant community composition
| Group | Common name | Symbol | Scientific name | Annual production (lb/acre) | Foliar cover (%) | |
|---|---|---|---|---|---|---|
|
Grass/Grasslike
|
||||||
| 1 | Perennial Grasses | 210–450 | ||||
| mountain brome | BRMA4 | Bromus marginatus | 60–144 | – | ||
| slender wheatgrass | ELTR7 | Elymus trachycaulus | 60–144 | – | ||
| muttongrass | POFE | Poa fendleriana | 30–54 | – | ||
| thickspike wheatgrass | ELLA3 | Elymus lanceolatus | 30–54 | – | ||
| sedge | CAREX | Carex | 0–5 | – | ||
|
Forb
|
||||||
| 2 | Perennial | 60–120 | ||||
| nettleleaf giant hyssop | AGUR | Agastache urticifolia | 6–30 | – | ||
| western sweetroot | OSOC | Osmorhiza occidentalis | 6–30 | – | ||
| ragwort | SENEC | Senecio | 6–30 | – | ||
| Fendler's meadow-rue | THFE | Thalictrum fendleri | 6–30 | – | ||
| violet | VIOLA | Viola | 0–3 | – | ||
| larkspur | DELPH | Delphinium | 0–3 | – | ||
| lupine | LUPIN | Lupinus | 0–3 | – | ||
|
Shrub/Vine
|
||||||
| 3 | Primary Shrubs | 60–108 | ||||
| mountain big sagebrush | ARTRV | Artemisia tridentata ssp. vaseyana | 30–54 | – | ||
| mountain snowberry | SYOR2 | Symphoricarpos oreophilus | 30–54 | – | ||
| 4 | Secondary Shrubs | 6–30 | ||||
| currant | RIBES | Ribes | 3–15 | – | ||
| blue elderberry | SANIC5 | Sambucus nigra ssp. cerulea | 0–6 | – | ||
| creeping barberry | MARE11 | Mahonia repens | 0–6 | – | ||
| rubber rabbitbrush | ERNA10 | Ericameria nauseosa | 0–3 | – | ||
|
Tree
|
||||||
| 5 | Deciduous | 30–54 | ||||
Interpretations
Animal community
Livestock Interpretations:
This site has value for livestock grazing because of its shade and quality of forage that is consistently found in the aspen understory. Adaptive management will be necessary to successfully establish aspen suckers and retain biodiversity. Grazing management considerations include timing, intensity and duration of grazing. Early season grazing may cause more intense soil compaction, thereby limiting growth in many species. Domestic livestock consume aspen with increasing pressure through summer and early fall as preferred forage decreases in volume and nutritional quality (DeByle 1985, Fitzgerald et al 1986).
Mountain brome increases with grazing (Leege et al. 1981). A study by Mueggler (1967), found that with clipping, mountain brome increased in herbage production when clipped in June. When clipped in July mountain brome increased due to reduced competition from forb species. The study also found that after three successive years of clipping mountain brome started to show adverse effects. Mountain brome is ranked as highly valuable as elk winter forage (Kufeld 1973).
Slender wheatgrass is a perennial bunchgrass that tends to be short lived, however it spreads well by natural reseeding (Monsen et al. 2004). It is widely used in restoration seedings (Monsen et al. 2004). Slender wheatgrass tends to persists for a longer time than other perennial grasses when subjected to heavy grazing (Monsen et al. 1996, Monsen et al. 2004). Slender wheatgrass is palatable and nutritious for livestock.
Wildlife Interpretations:
This site provides valuable habitat for a variety of wildlife species. Domestic livestock, wild ungulates, rodents and hares utilize aspen stands and can have a measurable impact. A study by Krebill (1972) found that the majority of aspen decline within their study area was due to a combination of pathogenic fungi and insects which invade aspen trees damaged by big game (Krebill 1972). Browsing during the sapling stage reduces aspen growth, vigor and numbers (DeByle and Winokur 1985). Heavy browsing on aspen suckers may result in lower clone vigor to the point that suckering no longer takes place. Browsing pressure may allow aspen to regenerate but prevent the development of trees, and the aspen will grow instead as a dense shrub (Bradley et al. 1992). Because aspen stands are grazed by cattle and/or sheep and also have a significant population of wild ungulates, grazing management and game management are important for the health of aspen communities. Removal of forage, as well as disruption of nesting site cover, can have negative effects on both large and small mammals (Kie et al 1991).
Quaking aspen is important forage for large mammals. Elk (Alces alces) browse the bark, branches and sprouts of quaking aspen year-round throughout the West (Beck and Peek 2000, DeByle 1979, Howard 1996). Mule deer (Odocoileus hemionus) use quaking aspen year round especially if winters are mild, browsing leaves, buds, twigs, bark, and sprouts. New growth, after burns or clearcuts, are readily consumed by mule deer (Robin 2013). Moose (Alces americanus) occasionally occur in Nevada but will feed on the bark of quaking aspen in winter, the saplings in spring, and leaves and branches the rest of the year (Sheppard at al. 2006). Black bear (Ursus americanus), will eat stems and leaves of quaking aspen; however, forbs and other plants found in quaking aspen understory are preferred (Ulev 2007, Wildlife Action Plan 2012).
Several lagamorphs use quaking aspen habitat. Although aspen groves are at elevations where desert cottontail (Sylvilagus audubonii) are not normally found; desert cottontail may use aspen habitat where aspen groves occur at lower elevation with sagebrush and shrubland (DeByle 1979). Snowshoe hares (Lepus americanus) feed on quaking aspen in summer and spring and snowshoe hares will continue to use quaking habitat year round, particularly if the there is substantial conifer components provided (Debyle 1979). A threatened species, the American Pika (Ochotona princeps) will utilize quaking aspen stands in higher elevation habitat and have been documented to feed on quaking aspen buds, twigs, and bark (Wildlife Action Plan 2012, Howard 1996).
Rodents utilize aspen habitat for food and cover. Pocket gophers, (Thomomys monticola) a fossorial rodent favor quaking aspen stands (Linzey and Hammerson 2008). Aspen soils rarely freeze which are ideal for borrowing pocket gophers. Forbs and aspen sprouts also provide forage in the spring and summer (DeByle 1979). Rodents including deer mice (Peromyscous maniculatus) and least chipmunks (Tamias minimus) occupy quaking aspen habitat (Debyle 1979). The deer mouse was trapped more than any other rodent, consistently throughout several years, in quaking aspen stands according to Andersen et al. (1980). The least chipmunk has been trapped at near equal density as the deer mouse in aspen habitat (DeByle 1979, Anderson et al. 1980). The Inyo shrew (Sorex tenellus), Merriam’s shrew (Sorex merriami), montane shrew (Sorex monticolus), and Western jumping mouse (Zapus princeps) use the shrub and herbaceous cover within quaking aspen habitat for foraging and cover (Wildlife Action Plan 2012). The flying squirrel (Glaucomys sabrinus), although rarely seen because of its nocturnal habit, is estimated to be one of the most common mammal species found in aspen type forests (Debyle 1979). Larger rodents, such as the North American porcupine (Erethizon dorsatum) will eat quaking aspen in winter and spring months. In winter, porcupine eat the smooth outer bark of the upper trunk and branches, in spring they eat the buds and twigs (Howard 1996, Debyle 1979).
Beaver (Castor canadensis) use a large amount of aspen for building material to construct their dams. It is estimated that as many as 200 quaking aspen stems are required to support one beaver for a 1-year period. Beaver prefer the inner bark of aspen to that of other trees as food (Lanner 1984). They will consume the leaves, bark, twigs, and any diameters of quaking aspen branches (Innes 2013). Previous research has estimated that an individual beaver consumes 2 to 4 pounds (1-2 kg) of quaking aspen bark daily (DeByle 1985).
Quaking aspen provide feed and cover for a variety of bird species in Nevada. The northern goshawk (Accipiter gentilis) and flammulated owl (Psiloscops flammeolus) use mature overstory for nesting (Nevada Wildlife Action Plan 2012). Bird species including orange-crowned and yellow-rumped warblers (Vermivora celata and Dendroica coronata, respectively), broad-tailed hummingbirds (Selasphorus platycercus), robins (Turdus migratorius), house wrens (Troglodytes aedon), pewees (Contopus sordidulus), juncos (Junco hyemalis), and thrushes (Catharus ustulatus) nest and forage aspen stands. Dead trees are used by downy woodpeckers (Picoides pubescens), flickers (Colaptes auratus) and Lewis’s woodpeckers (Melanerpes lewis). (Lanner 1984, Wildlife Action Plan 2012). Birds such as the mountain bluebird (Sialia currucoides), tree swallow (Tachycineta bicolor), pine siskin, (Spinus pinus), and black-headed grosbeak (Pheucticus melanocephalus) can be found at the edges of aspen communities (Innes 2013 and references therein). Even duck species, including, Wood duck (Aix sponsa), common and barrow’s goldeneye (Bucephala clangula and Bucephala islandica, respectively), bufflehead (Bucephala albeola), hooded and common merganser (Lophodytes cucullatus and Mergus merganserall, respectively) utilize aspen habitat (DeByle et al. 1985). Dusky grouse (Dendragapus obscurus), sooty grouse (Dendragapus fuliginosus), mountain quail (Oreotoryz pictus) and rufous hummingbird (Selasphorus rufus) utilize the shrub and herbaceous cover provided by quaking aspen forests (Wildlife Action Plan 2012).
Several bat species occur within subalpine habitat, adding to the community’s diversity. The fringed myotis (Myotis thysanodes), Long-eared myotis (myotis evotis), hoary bat (Lasiurus cinereus), Silver-haired bat (Lasionycteris noctivagans), little brown myotis (Myotis lucifugus), and western small-footed myotis (Myotis ciliolabrum) all are documented as occurring in quaking aspen forests and meadows above 9000 feet (Keinath 2003, Arroyo-Calbrales and Alvares-Castneda 2008, Warner and Czaplewski 1984, Armstrong 2007, Sullivan 2009, Great Basin National Park, Listing Sensitive and Extirpated Species 2006, Wildlife Action Plan 2012).
Habitat distribution of reptiles and amphibians is not as widely studied as other animals and few reptiles and amphibians are found at such elevations where quaking aspen trees occur. However; the Columbia spotted frog (Rana luteiventris) and northern rubber boa (Charina bottae) favor downed quaking aspen trees as well as stored ground moisture maintained from dead, decomposing logs (Wildlife Action Plan 2012).
Threats and management
Quaking aspen habitat is enhanced by periodic disturbance events and fire suppression, improper livestock grazing and browsing by big game species threaten aspen communities throughout Nevada (Wildlife Action Plan 2012). Several fungi species cause the formation of large caners on aspen trunks, roots and spots on leaves. The fungus Marssonina leaf-spot causes particular damage to the trees, leaving brown leaves on quaking aspen mid-summer throughout large portions of their habitat (Lanner 1984).
Hydrological functions
Permeability is slow to moderate. Runoff is medium to high. Hydrologic soil group is B. There is a shift in water use and yield of conifer-dominated versus aspen dominated forest with current thinking that coniferous forest use more water, i.e. a net reduction in runoff and recharge. As aspen stands are managed to favor conifers, water yield is diminished compared to an aspen stand.
Recreational uses
Aesthetic value is derived from the rich hues and textures of the aspen trees, particularly in the fall. The diverse flora and fauna, and the colorful wildflowers in the summer enhance the beauty of this site. The site offers rewarding opportunities to photographers and for nature study. It has high value for hunting, camping, picnicking, cross country skiing and family wood gathering. Management of the aspen woodland should include small, irregularly shaped clearcuts that blend into the natural landscape. Harvesting plans should include a mix of even-aged aspen patches in all size classes. Aspen fits well into management for dispersed recreation activities, but does not tolerate concentrated use, as found in established campgrounds. Encouraging concentrated recreation or developing campgrounds within aspen stands can lead to serious damage, including carving on trees, vandalism, destruction or removal of young suckers and trampling and disturbance of the soil.
Wood products
Historically quaking aspen has been used for mine props, posts, bridge planking, flooring, furniture and fuelwood. This tree has a considerable potential for increased utilization. It makes excellent pulp, excelsior, door corestock, paper, particleboard, matchsticks, structural flakeboard, lumber products and boxwood.
PRODUCTIVE CAPACITY
This site has a low site quality for quaking aspen production. Site index ranges from 35 to 40.
Productivity Class: 1
CMAI*: 12 to 16 cu ft/ac/yr;
0.7 to 1.1 cu m/hr/yr.
*CMAI: is the culmination of mean annual increment or highest average growth rate of the stand in the units specified.
Fuelwood Production: 3 to 8 cords per acre. There are about 203,000 gross British Thermal Units (BTUs) heat content per cubic foot of quaking aspen wood. Firewood is commonly measured by cord, or a stacked unit equivalent to 128 cubic feet. Solid wood volume in a cord varies, but assuming an average of 75 cubic feet of solid wood per cord, there are about 15 million BTUs of heat value in a cord of quaking aspen.
Saw timber: 100 to 200 board-feet per acre.
MANAGEMENT GUIDES AND INTERPRETATIONS
1. LIMITATIONS AND CONSIDERATIONS
a. Potential for sheet and rill erosion is moderate to severe depending on slope.
b. Moderate to severe equipment limitations on steeper slopes.
c. Proper spacing is the key to a well managed, multiple use aspen woodland.
d. Cut residual, unmerchantable, trees to stimulate maximum sucker regeneration and rapid development of a replacement stand.
e. Soil compaction and erosion hazards are greatest if logging is done with heavy equipment when soils are saturated in late spring. Logging at this time is most damaging to aspen roots, which can reduce suckering. Root carbohydrate reserves are lowest in spring and harvesting at this time can reduce sprouting.
2. ESSENTIAL REQUIREMENTS
a. Adequately protect from high intensity wildfire.
b. Protect soils from accelerated erosion.
c. Apply proper grazing management.
3. SILVICULTURAL PRACTICES
a. Harvest cut selectively or in small patches to enhance forage production.
1) Clearcutting - Clearcutting is appropriate when the primary management objective is sustained production of forest products, either sawtimber or fiber. Cutting submerchantable stems along with the merchantable ones will maximize sucker production, will minimize the presence of diseased or defective growing stock in the new stand, and will avoid suppression of the new crop by residual overstory stems.
2) Partial cutting - Partial cutting is feasible in uneven-aged stands where management objectives require vertical canopy diversity or retention of some overstory; partial cutting may result in enough sprouting to adequately regenerate stands. Extreme care is necessary to avoid injury to residual stems during logging.
3) Selective harvest - Selectively remove trees on suitable sites to enhance forage production and manage site reproduction.
b. Thinning - Ordinarily, only stands on sawtimber sites should be thinned. Precommercial thinning may be uneconomical. The low productivity of this site would not justify thinning costs.
c. Protection from disease - There are no proven forest stand treatments that successfully prevent or control disease in aspen. Maintenance of well-stocked stands, minimizing wounding of stems and control damaging agents, and harvesting at the proper rotation age are the best management recommendations that can be made today.
d. Protection from insects - Direct control of insects in aspen forests has not been practical. The environmental side-effects from chemical pesticide spraying usually has not been acceptable in the aspen ecosystem. Maintenance of a well-stocked stand and protection from wounding is the most practical method of coping with insects in the aspen forest.
e. Protection from mammals - Domestic livestock, wild ungulates, porcupines, rodents and hares utilize aspen as food and can have measurable impacts on some stands. Most animal damage can be prevented by careful husbandry of domestic livestock and by population control of wild game. Because most aspen stands are grazed by cattle and/or sheep and have a significant population of wild ungulates, grazing management and game management are important to aspen communities.
f. Fire management - Fire is a natural feature of the aspen ecosystem and is considered responsible for the abundance of aspen in the west, and for the even-aged structure of many stands. Without human intervention, fire appears to be necessary for the continued well-being of aspen on sites where natural degeneration of the clone occurs, or where insects or pests are especially harmful to the stand. Aspen forests do not readily burn. Fires in aspen generally are infrequent, spread slowly, are of low intensity, and are easy to control. In most areas, the conditions suitable for a fire are infrequent in aspen stands. Although aspen forests do not burn readily, aspen trees are extremely sensitive to fire. Even very light fires will kill aspen, because the bark is thin and green, and lacks protective corky layers. Even with adequate fuels, the flammability of adjacent grasslands may make prescribed burning risky. However, where fire can be used with reasonable safety, it is an inexpensive and effective way to naturally regenerate the aspen forest. Moderate intensity fire that kills most or all the overstory will stimulate adequate suckering and will have the least effect on subsequent sucker growth. If fire occurs at infrequent intervals and is moderately intense to kill most or all of the aspen overstory, most aspen sites will remain viable. Frequent fires may adversely affect site quality.
Other products
Common snowberry fruits were eaten fresh and also dried for winter use by Native Americans. Common snowberry was also used on hair as soap, and the fruits and leaves mashed and applied to cuts or skin sores as a poultice and to soothe sore, runny eyes. Tea from the bark was used as a remedy for tuberculosis and sexually transmitted diseases. A brew made from the entire plant was used as a physic tonic. Arrowshafts and pipestems were made from the stems. Native Americans used big sagebrush leaves and branches for medicinal teas, and the leaves as a fumigant. Bark was woven into mats, bags and clothing.
Other information
Quaking aspens are used to stabilize soil and watersheds. The trees produce abundant litter that contains more nitrogen, phosphorus, potash and calcium than leaf litter of most other hardwoods. The litter decays rapidly, forming nutrient-rich humus that may amount to 25 tons per acre (oven-dry basis). The humus reduces runoff and aids in percolation and recharge of ground water. Mountain brome is an excellent native bunchgrass for seeding alone or in mixtures in disturbed areas, including depleted rangelands, burned areas, roadways, mined lands, and degraded riparian zones. Slender wheatgrass is widely used for revegetating disturbed lands. Slender wheatgrass is a short-lived perennial with good seedling vigor. It germinates and establishes quickly when seeded making it a good choice for quick cover on disturbed sites. It persists long enough for other, slower developing species to establish. It is especially valuable for use in saline soils. It has been used for rehabilitating mine spoils, livestock ranges, and wildlife habitat and watershed areas.
Table 7. Representative site productivity
| Common name | Symbol | Site index low | Site index high | CMAI low | CMAI high | Age of CMAI | Site index curve code | Site index curve basis | Citation |
|---|---|---|---|---|---|---|---|---|---|
| POTRT | 35 | 40 | 12 | 16 | – | – | – |
Supporting information
Type locality
| Location 1: White Pine County, NV | |
|---|---|
| UTM zone | N |
| UTM northing | 4355077 |
| UTM easting | 705518 |
| Latitude | 39° 19′ 15″ |
| Longitude | 114° 36′ 57″ |
| General legal description | Berrycreek type location – in the Schell Creek Range, Humboldt-Toiyabe National Forest, about 1.5 miles southwest of South Schell Peak. This site is found in Elko and White Pine counties, Nevada. |
Other references
Airola, D. A. 1980. Northeast Interior Zone: Vol. III - Birds & Vol. IV - Mammals. U.S. Gov. Printing Off.: 1980 -- 690-082/26.
Baker, F. S. 1925. Aspen in the central Rocky Mountain region. U.S. Department of Agriculture, Bulletin 1291. Washington D.C. p. 47
Blaisdell, J. P. 1953. Ecological effects of planned burning of sagebrush-grass range on the upper Snake River Plains. Technical Bulletin 1075. US Dept. of Agriculture, Washington D.C. p. 39
Blaisdell, J. P., R. B. Murray, and E. D. McArthur. 1982. Managing intermountain rangelands-sagebrush-grass ranges. Gen. Tech. Rep. INT-134. U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station, Ogden, UT.
Bradley, A. F., N. V. Noste, and W. C. Fischer. 1992. Gen. Tech. Rep. INT-287: Fire ecology of forests and woodlands in Utah. . U.S. Department of Agriculture, Forest Service, Intermountain Research Station, Ogden, UT.
Bunting, S. C., B. M. Kilgore, and C. L. Bushey. 1987. Guidelines for prescribed burning sagebrush-grass rangelands in the northern Great Basin. US Department of Agriculture, Forest Service, Intermountain Research Station Ogden, UT, USA.
Caudle, D., J. DiBenedetto, M. Karl, H. Sanchez, and C. Talbot. 2013. Interagency Ecological Site Handbook for Rangelands. Available at: http://jornada.nmsu.edu/sites/jornada.nmsu.edu/files/InteragencyEcolSiteHandbook.pdf. Accessed 4 October 2013.
Chambers, J., B. Bradley, C. Brown, C. D’Antonio, M. Germino, J. Grace, S. Hardegree, R. Miller, and D. Pyke. 2013. Resilience to stress and disturbance, and resistance to Bromus tectorum L. invasion in cold desert shrublands of western North America. Ecosystems 17:1-16.
Crane, M. F. and W. C. Fischer. 1986. Fire ecology of the forest habitat types of central Idaho. p. 86.
Daubenmire, R. 1970. Steppe vegetation of Washington. Technical Bulletin 62. Washington State University, College of Agriculture, Washington Agriculture Experiment Station, Pullman, WA.
Daubenmire, R. 1975. Plant succession on abandoned fields, and fire influences in a steppe area in southeastern Washington. Northwest Science 49:36-48.
Davidson, Ross W., T. Hinds, F. Hawksworth. 1959. Decay of aspen in Colorado. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Station Paper 45. Fort Collins, Colo. p. 14
DeByle, N.V. 1985. Animal Impacts. In: DeByle, N. V., and R.P. Winokur, editors. Aspen: Ecology and Management in the Western United States. General Technical Report RM-119, Rocky Mtn For & Rng Exp Sta, FS, USDA. 223-232.
DeByle, N.V., P.J. Urness, and D.L. Blank. 1989. Forage Quality in Burned and Un-burned Aspen Communities. Research Paper INT-404. Inter. Res. Sta., FS, USDA.
Debyle, Norbert V., and Robert P. Winokur, editors. 1985. Aspen: ecology and management in the western United States. USDA Forest Service General Technical Report RM-119. Rocky Mountain Forest and Range Experiment Station, Fort Collins, Colo. p. 283
Debyle, N. V., C.D. Bevins, W.C. Fischer. 1987. Wildfire occurrence in aspen in the interior western United States. Western jounal of applied forestry 2: 73-76.
Dayton, W. 1937. Range Plant Handbook. USDA, Forest Service. Bull.
Fechner, G. H., and Barrows, J. S. 1976. Aspen stands as wildfire fuel breaks. Aspen bibliography. Paper 5029. http://digitalcommons.usu.edu/aspen_bib/5029
Fire Effects Information System (Online; http://www.fs.fed.us/database/feis/plants/).
Fitzgerald, R.D., R.J. Hudson, and A.W. Bailey. 1986. Grazing preferences of cattle in regenerating aspen forest. Jo. of Range Management. 39: 13-18.
Frey, B. R., V. J. Lieffers, E. H. Hogg, and S. M. Landhäusser. 2004. Predicting landscape patterns of aspen dieback: mechanisms and knowledge gaps. Canadian Journal of Forest Research 34:1379-1390.
Hallsten, G.P., Q.D. Skinner, A.A. Beetle. 1987. Grasses of Wyoming. 3d ed. Laramie: University of Wyoming, Agricultural Experiement Station. 432 p.
Houghton, J.G., C.M. Sakamoto, and R.O. Gifford. 1975. Nevada’s Weather and Climate, Special Publication 2. Nevada Bureau of Mines and Geology, Mackay School of Mines, University of Nevada, Reno, NV.
Kasworm, W. F., L. R. Irby, and H. B. I. Pac. 1984. Diets of ungulates using winter ranges in northcentral Montana. Journal of Range Management 37:67-71.
Kay, C. E. 1997. Is aspen doomed?. Aspen bibliography. Paper 1478.
http://digitalcommons.usu.edu/aspen_bib/1478
Kie, J.G., C.J. Evans, E.R. Loft, and J.W. Menke. 1991. Foraging behavior by mule deer: The influence of cattle grazing. Jo. of Wildlife Management. 55: 665-674.
Krebill, R. G. 1972. Mortality of aspen on the Gros Ventre elk winter range. Aspen Bibliography. Paper 5398. http://digitalcommons.usu.edu/aspen_bib/5398
Kufeld, R. C. 1973. Foods eaten by the Rocky Mountain Elk. Journal of Range Management 26:106-113.
Lanner, R.M. Trees of the Great Basin - A Natural History. University of Nevada Press, Reno, NV. 215 pp.
Leege, T. A., D. J. Herman, and B. Zamora. 1981. Effects of cattle grazing on mountain meadows in Idaho. Journal of Range Management 34:324-328.
Leege, T. A. and W. O. Hickey. 1971. Sprouting of northern Idaho shrubs after prescribed burning. The Journal of Wildlife Management 35:508-515.
Meinecke, E. P. 1929. Quaking aspen: A study in applied forest pathology. U.S. Department of Agriculture, Technical Bulletin 155. Washington D.C. p. 34
Merrill, E. H., H. Mayland, and J. Peek. 1982. Shrub responses after fire in an idaho ponderosa pine community. The Journal of Wildlife Management 46:496-502.
Miller, R. F. and E. K. Heyerdahl. 2008. Fine-scale variation of historical fire regimes in sagebrush-steppe and juniper woodland: an example from California, USA. International Journal of Wildland Fire 17:245-254.
Monsen, S. B., R. Stevens, S. C. Walker, and N. E. West. 1996. The competitive influence of seeded smooth brome (Bromus inermis) and intermediate wheatgrass (Thinopyron intermedium) within aspen-mountain brush communities of central Utah. In: Rangelands in a Sustainable Biosphere: Proceedings of the Fifth International Rangeland Congress, Salt Lake City, Utah, USA, 23-28 July, 1995. Volume 1.
Monsen, S. B., R. Stevens, and N. L. Shaw. 2004. Grasses. Pp. 295-424 In: S.B. Monsen, R. Stevens [eds.] Restoring Western Ranges and Wildlands, vol. 2. Gen. Tech. Rep. RMRS-GTR-136-vol-2. USDA: Forest Service, Rocky Mountain Research Station, Fort Collins, CO.
Mueggler, W. F. 1967. Response of mountain grassland vegetation to clipping in southwestern Montana. Ecology 48:942-949.
National Oceanic and Atmospheric Administration. 2004. The North American Monsoon. Reports to the Nation. National Weather Service, Climate Prediction Center. Available online: http://www.weather.gov/
Neuenschwander, L. 1980. Broadcast burning of sagebrush in the winter. Journal of Range Management 33:233-236.
Nimir, M. B. and G. F. Payne. 1978. Effects of spring burning on a mountain range. Journal of Range Management 31:259-263.
Noste, N. V. and C. L. Bushey. 1987. Fire response of shrubs of dry forest habitat types in Montana and Idaho. Gen. Tech. Rep. INT-239.
Potter, Donald A. 1998. Forested Communities of the Upper Montane in the Central and Southern Sierra Nevada. Gen. Tech. Rep. PSW-GTR-169. Albany, CA: Pacific Southwest Research
Station, Forest Service, U.S. Department of Agriculture. 319 p.
Strand, Eva K., L. A. Vierling, S. C. Bunting, P. E. Gessler. 2009. Quantifying successional rates in western aspen woodlands: Current conditions, future predictions. Forest Ecology and Management 257: 1705-1715.
Stringham, T.K., P. Novak-Echenique, P. Blackburn, C. Coombs, D. Snyder and A. Wartgow. 2015. Final Report for USDA Ecological Site Description State-and-Transition Models, Major Land Resource Area 28A and 28B Nevada. University of Nevada Reno, Nevada Agricultural Experiment Station Research Report 2015-01. p. 1524.
Tilley, D. J., D. Ogle, L. St. John, L. Holzworth, W. Crowder, and M. Majerus. 2004. Mountain Brome. USDA NRCS plant guide. USDA NRCS Plant Materials Center. USDA NRCS Idaho State Office, Idaho. p. 5
Tisdale, E. W. and M. Hironaka. 1981. The sagebrush-grass region: A review of the ecological literature. University of Idaho, Forest, Wildlife and Range Experiment Station.
USDA-NRCS Plants Database (Online; http://www.plants.usda.gov).
USDA-NRCS. National Forestry Manual - Part 537. Washington, D.C.
Vose, J. M. and A. S. White. 1991. Biomass response mechanisms of understory species the first year after prescribed burning in an Arizona ponderosa-pine community. Forest Ecology and Management 40:175-187.
Whitesides, C.J. 2012. The Effects of Fossorial Mammals on Alpine Treeline Dynamics in the American West. Dissertation. digital.library.txstate.edu.
Wildlife Action Plan 2012. Nevada Wildlife Action Plan. Nevada Department of Wildlife, Reno, NV.
Wright, H. A. 1971. Why squirreltail Is more tolerant to burning than needle-and-thread. Journal of Range Management 24:277-284.
Wright, H. A. 1985. Effects of fire on grasses and forbs in sagebrush-grass communities. Pages 12-21 In K. Sanders, J. Durham [eds.] Rangeland Fire Effects; A Symposium: Boise, ID, USDI-BLM.
Young, R. P. 1983. Fire as a vegetation management tool in rangelands of the intermountain region. In S.B. Monsen, N. Shaw [eds.] Proceedings: Managing intermountain rangelands - improvement of range and wildlife habitats Gen. Tech. Rep. INT-GTR-157. U.S. Department of Agriculture, Forest Service. P. 18-31.
Ziegenhagen, L. L. 2003. Shrub reestablishment following fire in the mountain big sagebrush (Artemisia tridentata Nutt. ssp. vaseyana (Rydb.) Beetle) alliance. Thesis. Oregon State University.
Ziegenhagen, L. L. and R. F. Miller. 2009. Postfire recovery of two shrubs in the interiors of large burns in the Intermountain West, USA. Western North American Naturalist 69:195-205.
Contributors
RK/GKB
T.Stringham/P.Novak-Echenique
Rangeland health reference sheet
Interpreting Indicators of Rangeland Health is a qualitative assessment protocol used to determine ecosystem condition based on benchmark characteristics described in the Reference Sheet. A suite of 17 (or more) indicators are typically considered in an assessment. The ecological site(s) representative of an assessment location must be known prior to applying the protocol and must be verified based on soils and climate. Current plant community cannot be used to identify the ecological site.
| Author(s)/participant(s) | |
|---|---|
| Contact for lead author | |
| Date | |
| Approved by | |
| Approval date | |
| Composition (Indicators 10 and 12) based on | Annual Production |
Indicators
-
Number and extent of rills:
-
Presence of water flow patterns:
-
Number and height of erosional pedestals or terracettes:
-
Bare ground from Ecological Site Description or other studies (rock, litter, lichen, moss, plant canopy are not bare ground):
-
Number of gullies and erosion associated with gullies:
-
Extent of wind scoured, blowouts and/or depositional areas:
-
Amount of litter movement (describe size and distance expected to travel):
-
Soil surface (top few mm) resistance to erosion (stability values are averages - most sites will show a range of values):
-
Soil surface structure and SOM content (include type of structure and A-horizon color and thickness):
-
Effect of community phase composition (relative proportion of different functional groups) and spatial distribution on infiltration and runoff:
-
Presence and thickness of compaction layer (usually none; describe soil profile features which may be mistaken for compaction on this site):
-
Functional/Structural Groups (list in order of descending dominance by above-ground annual-production or live foliar cover using symbols: >>, >, = to indicate much greater than, greater than, and equal to):
Dominant:
Sub-dominant:
Other:
Additional:
-
Amount of plant mortality and decadence (include which functional groups are expected to show mortality or decadence):
-
Average percent litter cover (%) and depth ( in):
-
Expected annual annual-production (this is TOTAL above-ground annual-production, not just forage annual-production):
-
Potential invasive (including noxious) species (native and non-native). List species which BOTH characterize degraded states and have the potential to become a dominant or co-dominant species on the ecological site if their future establishment and growth is not actively controlled by management interventions. Species that become dominant for only one to several years (e.g., short-term response to drought or wildfire) are not invasive plants. Note that unlike other indicators, we are describing what is NOT expected in the reference state for the ecological site:
-
Perennial plant reproductive capability:
Print Options
Sections
Font
Other
The Ecosystem Dynamics Interpretive Tool is an information system framework developed by the USDA-ARS Jornada Experimental Range, USDA Natural Resources Conservation Service, and New Mexico State University.
Click on box and path labels to scroll to the respective text.