
Natural Resources
Conservation Service
Ecological site R028BY034NV
MOUNTAIN RIDGE 12-14 P.Z.
Accessed: 11/24/2024
General information
Provisional. A provisional ecological site description has undergone quality control and quality assurance review. It contains a working state and transition model and enough information to identify the ecological site.
Figure 1. Mapped extent
Areas shown in blue indicate the maximum mapped extent of this ecological site. Other ecological sites likely occur within the highlighted areas. It is also possible for this ecological site to occur outside of highlighted areas if detailed soil survey has not been completed or recently updated.
MLRA notes
Major Land Resource Area (MLRA): 028B–Central Nevada Basin and Range
MLRA 28B occurs entirely in Nevada and comprises about 23,555 square miles (61,035 square kilometers). More than nine-tenths of this MLRA is federally owned. This area is in the Great Basin Section of the Basin and Range Province of the Intermontane Plateaus. It is an area of nearly level, aggraded desert basins and valleys between a series of mountain ranges trending north to south. The basins are bordered by long, gently sloping to strongly sloping alluvial fans. The mountains are uplifted fault blocks with steep sideslopes. Many of the valleys are closed basins containing sinks or playas. Elevation ranges from 4,900 to 6,550 feet (1,495 to 1,995 meters) in the valleys and basins and from 6,550 to 11,900 feet (1,995 to 3,630 meters) in the mountains.
The mountains in the southern half are dominated by andesite and basalt rocks that were formed in the Miocene and Oligocene. Paleozoic and older carbonate rocks are prominent in the mountains to the north. Scattered outcrops of older Tertiary intrusives and very young tuffaceous sediments are throughout this area. The valleys consist mostly of alluvial fill, but lake deposits are at the lowest elevations in the closed basins. The alluvial valley fill consists of cobbles, gravel, and coarse sand near the mountains in the apex of the alluvial fans. Sands, silts, and clays are on the distal ends of the fans.
The average annual precipitation ranges from 4 to 12 inches (100 to 305 millimeters) in most areas on the valley floors. Average annual precipitation in the mountains ranges from 8 to 36 inches (205 to 915 millimeters) depending on elevation. The driest period is from midsummer to midautumn. The average annual temperature is 34 to 52 degrees F (1 to 11 degrees C). The freeze-free period averages 125 days and ranges from 80 to 170 days, decreasing in length with elevation.
The dominant soil orders in this MLRA are Aridisols, Entisols, and Mollisols. The soils in the area dominantly have a mesic soil temperature regime, an aridic or xeric soil moisture regime, and mixed or carbonatic mineralogy. They generally are well drained, loamy or loamyskeletal, and shallow to very deep.
Nevada’s climate is predominantly arid, with large daily ranges of temperature, infrequent severe storms and heavy snowfall in the higher mountains. Three basic geographical factors largely influence Nevada’s climate: continentality, latitude, and elevation. The strong continental effect is expressed in the form of both dryness and large temperature variations. Nevada lies on the eastern, lee side of the Sierra Nevada Range, a massive mountain barrier that markedly influences the climate of the State. The prevailing winds are from the west, and as the warm moist air from the Pacific Ocean ascend the western slopes of the Sierra Range, the air cools, condensation occurs and most of the moisture falls as precipitation. As the air descends the eastern slope, it is warmed by compression, and very little precipitation occurs. The effects of this mountain barrier are felt not only in the West but throughout the state, as a result the lowlands of Nevada are largely desert or steppes.
The temperature regime is also affected by the blocking of the inland-moving maritime air. Nevada sheltered from maritime winds, has a continental climate with well-developed seasons and the terrain responds quickly to changes in solar heating. Nevada lies within the midlatitude belt of prevailing westerly winds which occur most of the year. These winds bring frequent changes in weather during the late fall, winter and spring months, when most of the precipitation occurs.
To the south of the mid-latitude westerlies, lies a zone of high pressure in subtropical latitudes, with a center over the Pacific Ocean. In the summer, this high-pressure belt shifts northward over the latitudes of Nevada, blocking storms from the ocean. The resulting weather is mostly clear and dry during the summer and early fall, with occasional thundershowers. The eastern portion of the state receives noteworthy summer thunderstorms generated from monsoonal moisture pushed up from the Gulf of California, known as the North American monsoon. The monsoon system peaks in August and by October the monsoon high over the Western U.S. begins to weaken and the precipitation retreats southward towards the tropics (NOAA 2004).
Ecological site concept
This site occurs on convex portions of mountains. Slopes gradients of 4 to 30 percent are typical. Elevations range from 6800 to 9000 feet.
Soils associated with this site are shallow, well drained, and formed in residuum and colluvium derived from mixed parent material. The soil profile has a mollic epipedon, an argillic and is skeletal with >35 percent rock fragments throughout. Soil moisture regime is xeric and the soil temperature regime is frigid.
The reference state is dominated by bluebunch wheatgrass, Thurber's needlegrass, low sagebrush and perennial mat-forming forbs. Black sagebrush may occur and is typically restricted to the ridge crest areas where soil depth is most limited. Production ranges from 100 to 350 pounds per acre.
Important abiotic factors contributing to the presence of this site include intense winds that inhibit snow accumulation and a convex landform shape, thus lowering the effective precipitation. The soils are shallow, have a high amount of gravels and cobbles armoring the surface and occuping plant growing space. The available water holding capacity is very low. Runoff is high to very high.
Associated sites
| R028BY015NV |
LOAMY SLOPE 12-16 P.Z. |
|---|---|
| R028BY030NV |
LOAMY 12-16 P.Z. |
| R028BY046NV |
GRAVELLY LOAM 12-14 P.Z. |
| R028BY087NV |
GRAVELLY CLAY 12-14 P.Z. |
Similar sites
| R028BY048NV |
CALCAREOUS MOUNTAIN RIDGE ARAR8 absent; soils from limestone parent material |
|---|---|
| R028BY027NV |
SHALLOW CALCAREOUS SLOPE 14+ P.Z. PSSPS dominant grass; ACTH7 absent; ARAR8 absent |
| R028BY039NV |
COBBLY CLAYPAN 12-14 P.Z. ARNO4 absent |
| R028BY037NV |
CLAYPAN 12-14 P.Z. ARNO4 absent; more productive site |
| R028BY038NV |
MOUNTAIN RIDGE 14+ P.Z. PSSPS dominant grass; POFE major grass; higher elevations |
Table 1. Dominant plant species
| Tree |
Not specified |
|---|---|
| Shrub |
(1) Artemisia arbuscula |
| Herbaceous |
(1) Pseudoroegneria spicata subsp. spicata |
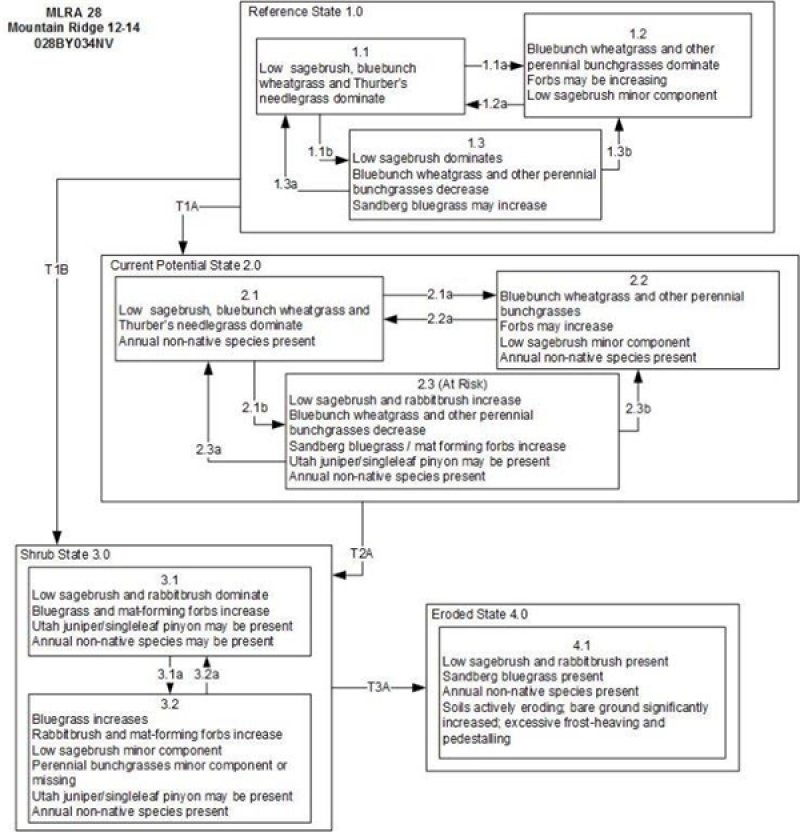



Click on box and path labels to scroll to the respective text.