
Natural Resources
Conservation Service
Ecological site FX052X01X165
Thin Claypan (Tcp) Dry Grassland
Last updated: 5/08/2025
Accessed: 03/24/2026
General information
Provisional. A provisional ecological site description has undergone quality control and quality assurance review. It contains a working state and transition model and enough information to identify the ecological site.
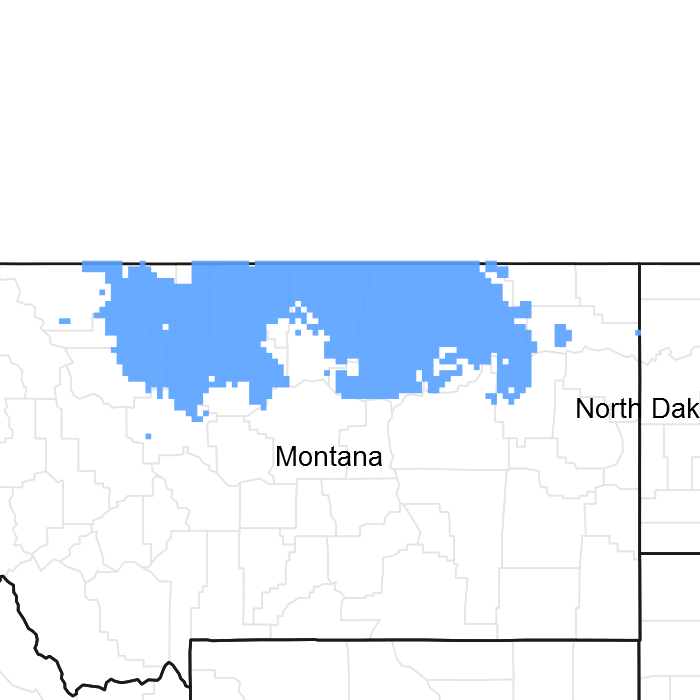
Figure 1. Mapped extent
Areas shown in blue indicate the maximum mapped extent of this ecological site. Other ecological sites likely occur within the highlighted areas. It is also possible for this ecological site to occur outside of highlighted areas if detailed soil survey has not been completed or recently updated.
MLRA notes
Major Land Resource Area (MLRA): 052X–Brown Glaciated Plains
The Brown Glaciated Plains, MLRA 52, is an expansive, agriculturally and ecologically significant area consisting of around 14.5 million acres that stretches across 350 miles from east to west, encompassing portions of 15 counties in north-central Montana. This region represents the southwestern limit of the Laurentide Ice Sheet and is considered to be the driest and westernmost area within the vast network of glacially-derived prairie pothole landforms of the northern Great Plains. The elevation ranges from 2,000 feet (610 meters) to 4,600 feet (1,400 meters).
Soils are primarily Mollisols but Entisols, Inceptisols, Alfisols and Vertisols are also common. Till from continental glaciation is the predominant parent material, but alluvium and bedrock are also frequently encountered. Till deposits are typically less than 50 feet thick and, in some areas, glacially deformed bedrock can be found at or near the soil surface (Soller, 2001). Underlying sedimentary bedrock largely consisting of Cretaceous shale, sandstone and mudstone (Vuke et al., 2007) is commonly exposed on hillslopes, particularly along drainageways. Significant alluvial deposits occur along glacial outwash channels and major drainages, which include portions of the Missouri, Teton, Marias, Milk and Frenchman Rivers. Large glacial lakes, particularly in the western half of the MLRA, deposited clayey and silty lacustrine sediments (Fullerton et al., 2013).
Much of the western portion of this MLRA was glaciated towards the end of the Wisconsin age with the maximum glacial extent occurring approximately 20,000 years ago (Fullerton et al., 2004). The result is a geologically young landscape that is predominantly a level till plain interspersed with lake plains and dominated by soils in the Mollisol and Vertisol orders. These soils are very productive and generally are well-suited to dryland farming. Much of this area is aridic-ustic and crop-fallow dryland wheat farming is the predominant land use, with rangeland typically being found on steep hillslopes along drainages.
Rangeland, much of it is native mixed grass prairie, increases in abundance in the eastern half of the MLRA. The Wisconsin-age till in the north-central part of this area typically formed large disintegration moraines with steep slopes and numerous poorly drained potholes. A large portion of Wisconsin-age till that is found on this type of terrain, which typically be optimal for farming, has large amounts of less-suitable sodium-affected Natrustalfs. Significant portions of Blaine, Phillips, and Valley Counties were glaciated approximately 150,000 years ago during the Illinoisan age and, due to erosion and dissection of the landscape, much of these areas have steeper slopes and more exposed bedrock than areas glaciated during the Wisconsin age (Fullerton et al., 1986).
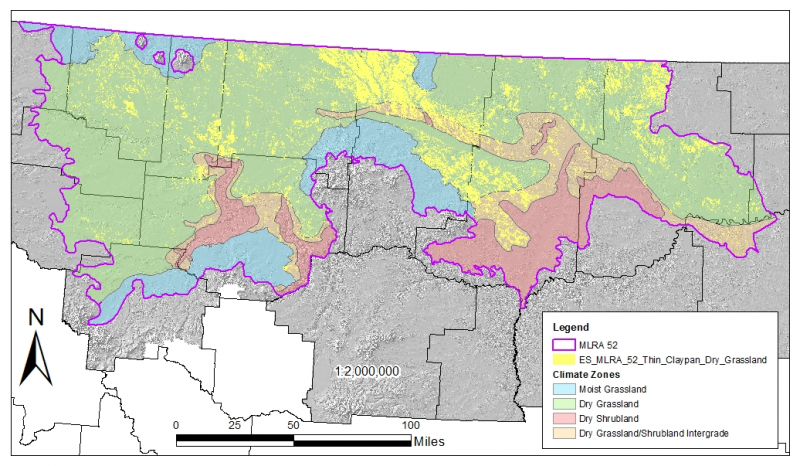
While much of the rangeland in the aridic ustic portion of MLRA 52 is classified as belonging to the ‘dry grassland’ climatic zone, sites in portions of southern MLRA 52 may belong to the ‘dry shrubland’ climatic zone. The dry shrubland zone represents the northernmost extent of big sagebrush (Artemisia tridentata) steppe on the Great Plains. As similar soils occur in both the southern and northern portions of the MLRA, it is currently hypothesized that climate is the primary driving factor affecting big sagebrush distribution in this area, but the precise factors have so far proven to be elusive and are, for the time of this writing, not yet fully understood.
Sizable tracts of largely unbroken rangeland in the eastern half of the MLRA and adjacent southern Saskatchewan are home to the Northern Montana Population of Greater Sage Grouse (Centrocercus urophasianus) and large portions of this area are considered to be a Priority Area for Conservation (PAC) by the U.S. Fish and Wildlife Service (U.S. Fish and Wildlife Service, 2013). This population is unique among sage grouse populations in the fact that many individuals overwinter in the big sagebrush steppe (dry shrubland) in the southern portion of the MLRA and then migrate to the northern portion of the MLRA which lacks big sagebrush (dry grassland) to spend rest of the year (Smith, 2013).
Areas of the till plain near the Bearspaw Mountains, Highwood Mountains, Sweetgrass Hills, and Rocky Mountain foothills are at higher elevation, receive higher amounts of precipitation, and have a typic-ustic moisture regime. These areas have significantly more rangeland production than the drier aridic-ustic portions of the MLRA and have enough moisture to produce crops annually as opposed to biennially in the drier areas. Ecological sites in this higher precipitation area are classified as the moist grassland climatic zone.
Classification relationships
NRCS Soil Geography Hierarchy
Land Resource Region: Northern Great Plains
Major Land Resource Area: 52
Climatic Zone: Dry Grassland
National Hierarchical Framework of Ecological Units (Cleland et al., 1997; Nesser et al., 1997; McNab et al., 2007)
• Domain: Dry
• Division: Temperate Steppe
• Province: Great Plains-Palouse Dry Steppe Province 331
• Section: Northwestern Glaciated Plains 331D
• Subsection: Montana Glaciated Plains 331Dh
• Landtype association/Landtype phase: N/A
National Vegetation Classification Standard (Federal Geographic Data Committee, 2008)
• Class: Mesomorphic Shrub and Herb Vegetation Class (2)
• Subclass: Temperate and Boreal Grassland and Shrubland Subclass (2.B)
• Formation: Temperate Grassland, Meadow, and Shrubland Formation (2.B.2)
• Division: Great Plains Grassland and Shrubland Division (2.b.2.Nb)
• Macrogroup: Hesperostipa comata – Pascopyrum smithii – Festuca hallii Grassland Macrogroup (2.B.2.Nb.2)
• Group: Pascopyrum smithii – Hesperostipa comata – Schizachyrium scoparium – Bouteloua spp. Mixedgrass Prairie Group (2.B.2.Nb.2.c)
• Alliance: Pascopyrum smithii – Nassella viridula Northwestern Great Plains Herbaceous Alliance
• Association: Pascopyrum smithii –Bouteloua gracilis – Carex filifolia Herbaceous Vegetation
EPA Ecoregions
• Level 1: Great Plains (9)
• Level 2: West-Central Semi-Arid Prairies (9.3)
• Level 3: Northwestern Glaciated Plains (42)
• Level 4: North Central Brown Glaciated Plains (42o) and Glaciated Northern Grasslands (42j)
Ecological site concept
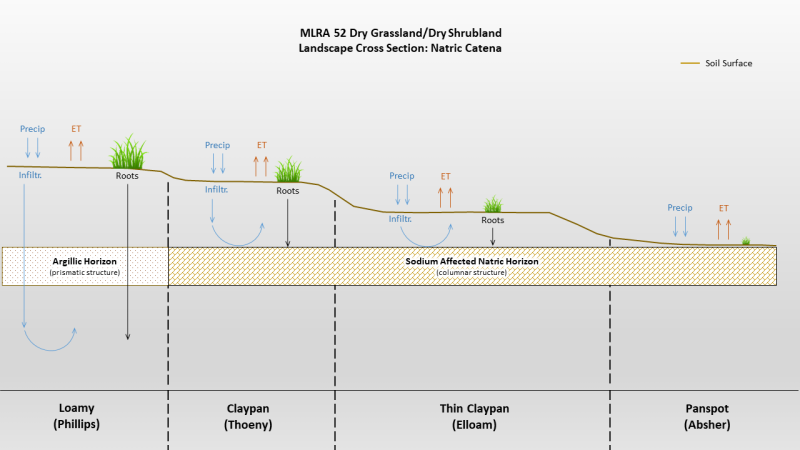
The Thin Claypan Dry Grassland ecological site occurs on till plains, moraines and fans, generally on slopes less than eight percent or less. The site is characterized by a dense root-restricting layer (evidenced by columnar structure) that is between one (1) and four (4) inches from the soil surface. The surface texture is typically loam over clay or clay loam. Vegetation is dominated by western wheatgrass (Pascopyrum smithii) and/or thickspike wheatgrass (Elymus lanceolatus). Less common grasses include plains reedgrass (Calamagrostis montanensis), blue grama (Bouteloua gracilis), prairie Junegrass (Koeleria macrantha) and Sandberg bluegrass (Poa secunda). Silver sagebrush is the most common shrub. This site is typically associated with Panspot and Claypan ecological sites. Figure 1 illustrates the distribution of this ecological site based on current data. This map is approximate, is not intended to be definitive, and may be subject to change. Field verification using the appropriate MLRA key is required for proper identification of this ecological site.
Associated sites
| FX052X01X006 |
Claypan (Cp) Dry Grassland Claypan is found on higher micro topography than Thin Claypan Dry Grassland, but lower than Loamy. |
|---|---|
| FX052X01X032 |
Loamy (Lo) Dry Grassland Loamy is found on the highest micro topography whereas Thin Claypan Dry Grassland is much lower. |
| FX052X01X145 |
Panspot (Pn) Dry Grassland Panspot is found on the lowest micro topography whereas Thin Claypan Dry Grassland occupies higher positions. |
Similar sites
| FX052X03X165 |
Thin Claypan (Tcp) Dry Shrubland Differs from Thin Claypan Dry Grassland in that annual temperatures are slightly warmer and site supports big sagebrush rather than silver sagebrush. |
|---|---|
| FX052X01X145 |
Panspot (Pn) Dry Grassland Differs from Thin Claypan Dry Grassland in that the root restricting layer (evidenced by columnar structure) is 1 inch or less from the surface. |
| FX052X01X006 |
Claypan (Cp) Dry Grassland Differs from Thin Claypan Dry Grassland in that the root restricting layer (evidenced by columnar structure) is greater than 4 inches to 10 inches below the soil surface. |
| FX052X01X032 |
Loamy (Lo) Dry Grassland Differs from Thin Claypan Dry Grassland in that the root restricting layer (evidenced by columnar structure) is either absent or greater than 10 inches below the soil surface. |
Table 1. Dominant plant species
| Tree |
Not specified |
|---|---|
| Shrub |
Not specified |
| Herbaceous |
(1) Pascopyrum smithii |
Legacy ID
R052XY165MT



Click on box and path labels to scroll to the respective text.
| T1A | - | prolonged drought, improper grazing, or a combination of these factors |
|---|---|---|
| T1B | - | The introduction of aggressive perennial grasses (mostly crested wheatgrass) |
| T1C | - | Conversion to annual cropland |
| R2A | - | range seeding, grazing land mechanical treatment, timely moisture, proper grazing management (management intensive and costly) |
| T2A | - | introduction of weedy species; combined with drought and/or improper grazing management |
| T2B | - | conversion to annual cropland |
| T3A | - | conversion to annual cropland |
| T4A | - | cessation of annual cropping |
| T5A | - | conversion to annual cropland |
State 1 submodel, plant communities
| 1.1A | - | drought, improper grazing management, multiple fires in close succession |
|---|---|---|
| 1.2A | - | timely moisture, proper grazing management |