
Natural Resources
Conservation Service
Ecological site R018XI102CA
Thermic Ultramafic Foothills Extremely High Magnesium Content (Ca:Mg Ratio Less Than 0.5)
Accessed: 04/10/2026
General information
Approved. An approved ecological site description has undergone quality control and quality assurance review. It contains a working state and transition model, enough information to identify the ecological site, and full documentation for all ecosystem states contained in the state and transition model.
Figure 1. Mapped extent
Areas shown in blue indicate the maximum mapped extent of this ecological site. Other ecological sites likely occur within the highlighted areas. It is also possible for this ecological site to occur outside of highlighted areas if detailed soil survey has not been completed or recently updated.
MLRA notes
Major Land Resource Area (MLRA): 018X–Sierra Nevada Foothills
Major Land Resource Area (MLRA) 18, Sierra Nevada Foothills is located entirely in California and runs north to south adjacent to and down-slope of the Sierra Nevada Mountains (MLRA 22A). MLRA 18 includes rolling to steep dissected hills and low mountains, with several very steep river valleys. Climate is distinctively Mediterranean (xeric soil moisture regime) with hot, dry summers, and relatively cool, wet winters. Most of the precipitation comes as rain; average annual precipitation ranges from 18 to 40 inches in most of the area (precipitation generally increases with elevation and from south to north). Geology is rather complex in this region; there were several volcanic flow and ashfall events, as well as tectonic uplift, during the past 25 million years that contributed to the current landscape. Due to extreme latitudinal differences in MLRA 18, Land Resource Units (LRUs) were designated to group the MLRA into similar land units.
LRU Description:
This LRU (designated XI) occurs over a significant proportion of Mariposa, Tuolumne, Calaveras, Amador, El Dorado, Placer, and Nevada Countiesand is located on moderate to steep mountains and hills in the Sierra Nevada Foothills. It also includes mesa formations from volcanic flows, where vernal pool habitats occur. Various geologies occur in this region: metavolcanics, granodiorite, slate, marble, argillite, and quartzite, as well as ultramafic bands to a limited and localized extent. Soil temperature regime is thermic and soil moisture regime is xeric. Elevation ranges between 300 and 2000 ft above sea level. Precipitation ranges from 18 to 42 inches annually. Most precipitation falls between the months of November and March in the form of rain. Dominant vegetation includes annual grasslands, blue oak (Quercus douglasii), interior live oak (Quercus wislizeni), chamise (Adenostoma fasciculatum), buckbrush (Ceanothus cuneatus), and foothill pine (Pinus sabiniana).
Classification relationships
This site is located within M261F, the Sierra Nevada Foothills Section, (McNab et al., 2007) of the National Hierarchical Framework of Ecological Units (Cleland et al., 1997), 261Ca, the Western Foothills Subsection.
Level III and Level IV ecoregions systems (Omernik, 1987, and EPA, 2011) are: Level III, Central California Foothills and Coastal Mountains and Level IV, Ecoregion 6b, Northern Sierran Foothills.
Ecological site concept
Ultramafic bedrock is characterized by its low Ca:Mg ratios, which range between 0.05 and 0.5 and high heavy metal accumulations and is thus toxic to many plants; it is usually associated with stunted growth or reduced productivity. The influence of soil chemistry is readily apparent by virtue of its influence on vegetation composition, production, and species distribution. The plant communities in this ecological site are strikingly different than the adjacent non-serpentinite derived soils, as would be expected in a serpentinite area (McGahan et al., 2009; Kruckenburg, 1984). Serpentinite and similar ultramafic geologies occur in several ophiolite sequences throughout the region and are known to differ widely in vegetative expression. This expression may vary from completely barren ground to chaparral (Lazarus et al., 2011), to altered species composition and conifer density in forest communities (e.g. coastal Oregon, Kruckenburg, 1984). Although, there seems to be very little difference in expression on this ecological site.
The vegetative expression in this ecological site consists of buckbrush chaparral with scattered Foothill pine and grassland, occurring in stark contrast to the adjacent non-serpentine blue oak- annual grasslands. Several less common and endemic plants occur within this ecological site. The production RV is about 238 lbs per acre and ranges between 100 and 750 lbs per acre, depending on previous year’s precipitation received.
Associated sites
| R018XI103CA |
Thermic Ultramafic Foothills Moderately High Magnesium Content (Ca:Mg Ratio 0.5 To 2) This site occurs on adjacent hillslopes with higher Ca:Mg ratios (roughly 0.5 to 2). Less toxic soils support chamise chaparral with yerba santa (Eriodictyon californicum), toyon (Heteromeles arbutifolia) and buckbrush (Ceanothus cuneatus) as secondary species. Occasional blue oaks (Quercus douglasii) have been found on this ecological site. |
|---|---|
| R018XI104CA |
Thermic Ultramafic North-Facing Steep Slopes This site occurs on adjacent steep (60-90% slopes) north-facing hillslopes. Shrubs like toyon (Heteromeles arbutifolia), manzanita (Arctostaphylos), and buckbrush (Ceanothus cuneatus)occur in understory of often dense California foothill pine stands. |
Table 1. Dominant plant species
| Tree |
(1) Pinus sabiniana |
|---|---|
| Shrub |
(1) Ceanothus cuneatus |
| Herbaceous |
Not specified |
Physiographic features
This site includes the ultramafic bands in Tuolumne and Calaveras Counties, CA which extend north into Amador, El Dorado, and Placer Counties; the most widespread area of this geologic feature occurs in the BLM Red Hills Area of Critical Environmental Concern (ACEC). Similar ecological sites occur in ultramafic bands extending among the entire Northern Sierra Foothill LRU, with some differences in soil chemistry and vegetative composition. It occurs on loamy to loamy-skeletal soils on footslopes and backslopes of all aspects and slope gradient ranges between 3 and 60%. Soils occur on elevations between 670 and 1,650 ft above sea level.
Table 2. Representative physiographic features
| Landforms |
(1)
Hill
|
|---|---|
| Flooding frequency | None |
| Ponding frequency | None |
| Elevation | 670 – 1,650 ft |
| Slope | 3 – 60% |
Climatic features
This ecological site is a found in a broad Mediterranean climatic region, with hot, dry summers and cool, wet winters. Mean annual precipitation is around 25 inches and ranges between 21 to 31 inches per year, mostly occurring between November and April in the form of rain. Mean annual air temperature ranges between 59 and 63 degrees F. The frost free period is 212 to 242 days and the freeze-free period is approximately 258 to 300 days.
Maximum and minimum monthly climate data for this ESD were generated by the Climate Summarizer
(http://www.nm.nrcs.usda.gov/technical/handbooks/nrph/Climate_Summarizer.xls) using data
from the following climate stations (equally weighted):
48353 Sonora Ranger Station
(Period of record = 1971 to 2000)
46172 New Melones Dam
(Period of record = 1992 to 2012)
44590 Knights Ferry 2 ESE
(Period of record = 1959 to 1977)
The data from multiple weather stations were combined to most accurately reflect the climatic conditions of this ecological site. The Knights Ferry weather station is closest to the ACEC site but is limited by the number of years and the time period data was collected. The New Melones Dam weather station is also in a nearby area, but is at slightly higher elevation, and precipitation was considerably higher. The Sonora Ranger Station had the most complete dataset (more than a century), however in order to match within the range of the other two climate stations the 1959 to 2012 records were used for precipitation and the 1971 to 2000 temperature averages were used for this document.
Table 3. Representative climatic features
| Frost-free period (average) | 227 days |
|---|---|
| Freeze-free period (average) | 279 days |
| Precipitation total (average) | 26 in |
Figure 2. Monthly precipitation range
Figure 3. Monthly average minimum and maximum temperature
Figure 4. Annual precipitation pattern
Figure 5. Annual average temperature pattern
Influencing water features
Soil features
The soils in this ecological site are formed from the colluvium and residuum of serpentinite and other ultramafic rock. Soils are shallow to moderately deep, and have loamy to loamy-skeletal (or clayey-skeletal) textures. The bedrock is a restrictive layer found between 10 and 39 inches of depth. Gravels (< 3 inch diameter) range between 0 to 30 % cover, while larger fragments (>= 3 inch diameter) are 14 to 32 % cover. Subsurface gravels range between 0 to 15 % and larger fragments occupy 0 to 31 % by volume. The soils in this ecological site are well drained and the permeability class ranges from slow to very slow.
The most commonly occurring soil component in the map units outlined below is Crimeahouse (loamy-skeletal, smectitic, thermic Mollic Haploxeralfs). This ecological site is also associated with major component of Hennekenot (loamy-skeletal, mixed, superactive, thermic Lithic Argixerolls); and minor component Delpiedra (loamy, magnesic, thermic shallow Mollic Haploxeralfs) soils. It is also associated with major component Henneke (clayey-skeletal, magnesic, thermic, Lithic Argixerolls).
The common characteristic shared by all soils in the Thermic Ultramafic Foothills is an unbalanced calcium:magnesium (Ca:Mg) ratio that is toxic to most plant species. The Delpiedra series is classified as having a magnesic mineralogy class (forty percent or more (by weight) magnesium silicate minerals such as serpentinite (antigorite, shrysotile, and lizardite) talc, olivines, Mg-rich pyroxenes, and/or Mg-rich amphiboles in the particle size control section. Henneke also has a magnesic mineralogy class and a shallow depth class to indurated bedrock. While Crimeahouse and Hennekenot do not have a magnesic mineralogy class, they still have very low Ca:Mg ratios in the subsurface horizons (0.1 to 0.2, and 0.2 to 0.57, respectively). The primary difference between Crimeahouse and Hennekenot is their depth class (moderately deep and shallow, respectively). Additionally, Crimeahouse has a darker epipedon than either Hennekenot or Delpiedra. The Delpiedra soils have loamy surface textures and belong to a shallow depth class (< 20 in). All components assigned to this ecological site have an argillic horizon, meaning that clay is being translocated to deeper horizons. Additionally, Henneke has a more fine texture (clayey-skeletal) than the other components above.
This ecological site is correlated with the following map units and soil components in MLRA 18:
20mn6; Crimeahouse-Delpiedra complex, 3 to 15 percent slopes; Crimeahouse;;55; Delpiedra;;35; Hennekenot;;10;
20mn7; Crimeahouse-Hennekenot-Delpiedra complex, 15 to 30 percent slopes; Crimeahouse;;50; Hennekenot;;30; Delpiedra;;20;
21zq2; Hennekenot-Crimeahouse complex, 30 to 60 percent slopes; Hennekenot;;50; Delpiedra;; 5;
hfzs; Henneke-Rock outcrop complex 5 to 50 percent slopes; Henneke;;65;
Table 4. Representative soil features
| Parent material |
(1)
Colluvium
–
ultramafic rock
|
|---|---|
| Surface texture |
(1) Loam (2) Stony loam (3) Stony clay loam |
| Drainage class | Well drained |
| Permeability class | Moderately slow to slow |
| Soil depth | 10 – 39 in |
| Surface fragment cover <=3" | 30% |
| Surface fragment cover >3" | 15% |
| Available water capacity (0-40in) |
0.6 – 4.1 in |
| Calcium carbonate equivalent (0-40in) |
Not specified |
| Electrical conductivity (0-40in) |
Not specified |
| Sodium adsorption ratio (0-40in) |
Not specified |
| Soil reaction (1:1 water) (0-40in) |
5.6 – 7.8 |
| Subsurface fragment volume <=3" (Depth not specified) |
14 – 32% |
| Subsurface fragment volume >3" (Depth not specified) |
31% |
Ecological dynamics
The main controlling factor in soils forming in ultramafic parent material is the chemical composition. The overwhelming abundance of extractable Mg at the cation exchange sites (at the expense of extractable Ca (Brooks, 1987)) prevents many plants from establishing. In addition to very low Ca:Mg ratios, serpentinite, dunite, and perioditite contain elevated levels of heavy metals (Woodruff et al., 2009, Ni Mn, etc.). The chemical composition is often heterogeneous in distribution (Alexander and Dushey, 2011). For instance, some ultramafic soils include barren pockets of highly toxic soil, where no plants grow (see Lazarus et al., 2011). In contrast, the ecological site described below seems to be relatively homogeneous in the effects of the ultramafic parent material on the vegetation.
Kruckeberg (1984) outlined vegetative response to ultramafic conditions: plants are 1) endemic to serpentine (restricted), 2) not restricted (e.g. local indicators), 3) indifferent to serpentine (Bodenvag), and 4) excluded from serpentine (e.g. blue oak). Buckbrush (Ceanothus cuneatus) falls under the second category of local indicator. Lazarus et al. (2011) found that buckbrush growing in pots from barren serpentinite were able to avoid accumulation of magnesium and other toxic elements to a greater degree than several restricted herbaceous plants. Buckbrush often grows in the understory of adjacent blue oak sites, yet at much lower densities than what is found in this ecological site. Given the species ability to avoid heavy metal accumulation, drought tolerance (Parsons et al., 1981), and N-fixation properties (Delwiche et al., 1965) buckbrush is the most common plant species in the Red Hills. Foothill pine, on the other hand is likely a Bodenvag or indifferent species. It also is found in adjacent ecological sites, yet its productivity is roughly equal in both ultramafic and non-ultramafic sites alike.
Ultramafic soils have been thought to be refuges for native endemics as well as perennial bunchgrasses (Kruckeburg, 1984; Huenneke, 1990; Gram et al., 2004). Opportunistic and/or targeted studies in this ecological site have documented endemic annual forbs such as the bilobed clarkia (Clarkia biloba spp. biloba) and Mariposa cryptantha (Cryptantha mariposae), and endemic perennial plants including Congdon’s lomatium (Lomatium congdonii), Rawhide Hill Onion (Allium tuolumnense), and Hernandez blue curls (Trichostema rubisepalum) (see (http://www.tuolumnewildflowers.com/resources/Red+Hills+2012+Observations.htm for a complete species list for this and similar ecological sites).
Disturbance dynamics
The two main historical disturbances in this region are grazing practices and fire. Livestock grazing has occurred for at least 200 years and has likely contributed to the spread of Mediterranean annual grasses such as Bromus and Avena genus (Jackson, 1985). The overall effect of cattle grazing for this ecological site is uncertain. The Red Hills of Tuolumne County, an area where this ecological site tends to dominate the landscape, has been excluded from cattle grazing since being listed as an area of critical environmental concern (ACEC) in 1993. Two active grazing allotments remain (Cranston, personal comm.). This makes comparison between grazed and ungrazed parcels a possibility. During a rapid assessment in 2013 in the Red Hills, it appeared that the livestock were not utilizing the upland where this ecological site exists, but rather they congregated near the riparian areas at the bottom of the drainages. Harrison et al. (2003) found that native species richness increased after grazing in a serpentine meadow. This ecological site is a chaparral dominated ecological site, therefore the effects would likely differ from grassland dominated systems. Furthermore, it must be recognized that domestic ungulates are not the only drivers of ecological dynamics. Native herbivores such as rabbits (Lepus spp.) and deer (Odocoileus spp.) utilize both the herbaceous and woody vegetation.
Contrastingly, fire has likely been a more significant shaping force over a much longer period than grazing. Native indigenous groups among the entire length of the Sierra Nevada Foothills practiced setting fires millennia before European establishment. The diverse array of reasons for burning, included hunting purposes, to maintain vegetation (clearing underbrush), and to improve crop yield (Stewart, 2002). Yet, only faint traces of Miwok natives’ inhabitation exist within the Red Hills ACEC, probably because of the scarcity of game and acorns (US Department of Interior, 2013). Nevertheless, it is likely that the native inhabitants played some role in maintaining the fire regime of this ecological site. The effects of fire in serpentinite dominated vegetation communities have been studied more extensively on the Central California Coastal Range (MLRA 15). For example, Safford and Harrison (2004) noted that serpentine chaparral differs in post-fire response than other chaparral types. Their study area showed longer intervals between fires in serpentine, in part because of less fuel loading due to relatively lower site productivity. The life history of buckbrush (League, 2005) suggests that a longer fire return interval may be beneficial to some plant species. For example, buckbrush is an obligate seeder that germinates following fire scarification (Biswell, 1963), but can remain viable in the seed bank for several years in absence of fire. Buckbrush plants begin senescing beyond 50 years (Biswell, 1963), in which time substantial fuel buildup will have occurred, raising the potential for wildfires. Modeling showed that another long-lived obligate-seeder (Ceanothus greggii) responded most favorably to a 35 to 50 year fire return interval (Regan et al., 2010). This interval is up to twice as long as the fire interval recorded in blue oak grasslands (McClaran, 1986). The last major fire recorded in the Red Hills ACEC was in 1996. Outside of the Red Hills, in the more narrow ultramafic bands, fire return intervals would likely be more variable and might be more similar to the 25 year common in surrounding blue oak woodlands.
Another anthropogenic disturbance occurring in chaparral is brush clearing, which oftentimes is coupled with a domestic livestock grazing regiment. Within the State of California, 1.9 million acres of hardwood and chaparral have been cleared (Bolsinger, 1988), since European settlement. It appears that the Red Hills have remained largely intact with only some localized brush clearing occurring and the only timber source, the California foothill pine, is a marginal tree species and seldom utilized. Nevertheless, the authors have found signs of recent clearing on private land adjacent to the ACEC. There are also unprotected stringers of this ecological site throughout Tuolumne and Calaveras Counties and into Amador to the north.
State and transition model
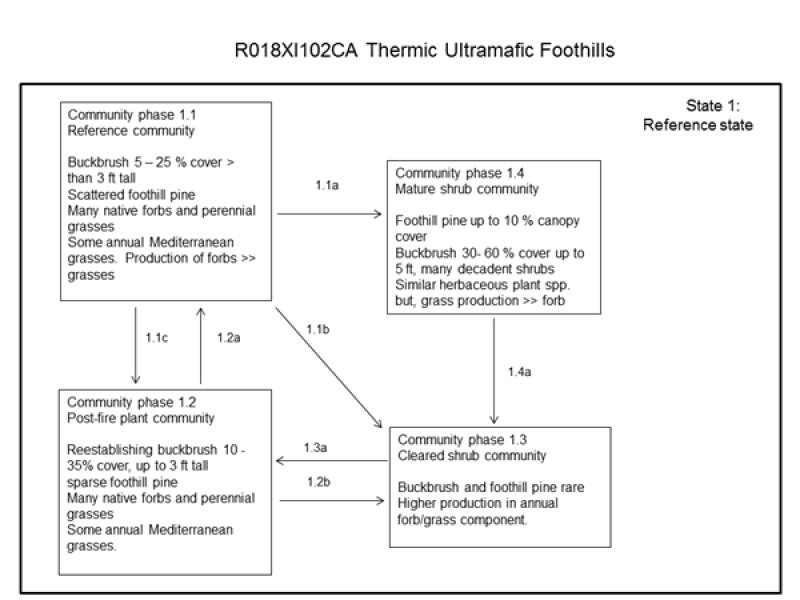
Figure 6. R018XI102CA STM
More interactive model formats are also available.
View Interactive Models
More interactive model formats are also available.
View Interactive Models
Click on state and transition labels to scroll to the respective text
Ecosystem states
State 1 submodel, plant communities
State 1
Reference State
State 1 represents the historic range of variability for this ecological site. This ecological site may be one of the few examples in California where historic plant communities are still observable. Ultramafic soils often offer a refuge to many native endemic herbaceous species because its unique soil chemistry is not amenable to the vast majority of plant species. Most of the endemic plants are still intact, despite the introduction of a number of annual grasses and forbs arriving after the discovery of the New World.
Community 1.1
Reference community

Figure 7. Community phase 1.1 during spring with buckbrush in flower; Photo: D. Evans, 2012.
This community phase is dominated by buckbrush (25-45% canopy cover) and scattered foothill pine (generally < 5 % canopy cover). Other shrubs may occasionally appear but are rare and occupy <1% of a site. Shrub interspaces are populated with grasses and forbs. Ground cover of herbaceous plants is discontinuous. Bare ground and surface fragments are common; larger bare surfaces (>10 square feet) may be composed of microsites of elevated toxicity (see Lazarus et. al., 2011). Fine woody fuels and course woody fuels are abundant, which would carry a fire during the summer or fall. Native forbs such as (but not limited to) annual agoseris (Agoseris heterophylla), cutleaf owl’s clover (Castilleja lacera), California goldfields (Lasthenia californica), and bluedicks (Dichelostemma capitana) are often present and flowering by March through early April. Just following those blooms, purple sanicle (Sanicula bipinnatifida), a tuberous perennial emerges. Early to mid-summer flowering plants include vinegarweed (Trichostema lanceolatum) and tripod buckwheat (Erigonum tripodum). Goldback fern (Pentagramma triangularis), usually found in trace amounts, tends to proliferate in some areas beneath shrub canopies. In addition to brome species, other non-native annual grasses such as nit grass (Gastridum pheloides), wildoat (Avena spp.) are often present. Additionally, the harsh chemical and climatic nature of these sites has acted as a refuge to a number of bunchgrasses, including (but not limited squirreltail (Elymus elymoides), melicgrass (Melica spp.), and California meadow barley (Hordeum brachyantherum spp. californicum.).
Figure 8. Annual production by plant type (representative values) or group (midpoint values)
Table 5. Annual production by plant type
| Plant type | Low (lb/acre) |
Representative value (lb/acre) |
High (lb/acre) |
|---|---|---|---|
| Forb | 17 | 171 | 262 |
| Grass/Grasslike | 6 | 33 | 172 |
| Tree | 0 | 40 | 165 |
| Shrub/Vine | 70 | 100 | 150 |
| Total | 93 | 344 | 749 |
Table 6. Soil surface cover
| Tree basal cover | 0% |
|---|---|
| Shrub/vine/liana basal cover | 2-3% |
| Grass/grasslike basal cover | 0-1% |
| Forb basal cover | 0-1% |
| Non-vascular plants | 7-35% |
| Biological crusts | 0% |
| Litter | 7-50% |
| Surface fragments >0.25" and <=3" | 2-36% |
| Surface fragments >3" | 0-15% |
| Bedrock | 0-3% |
| Water | 0% |
| Bare ground | 6-27% |
Table 7. Woody ground cover
| Downed wood, fine-small (<0.40" diameter; 1-hour fuels) | 0-10% |
|---|---|
| Downed wood, fine-medium (0.40-0.99" diameter; 10-hour fuels) | 2-5% |
| Downed wood, fine-large (1.00-2.99" diameter; 100-hour fuels) | 0-2% |
| Downed wood, coarse-small (3.00-8.99" diameter; 1,000-hour fuels) | 0-1% |
| Downed wood, coarse-large (>9.00" diameter; 10,000-hour fuels) | 0-2% |
| Tree snags** (hard***) | – |
| Tree snags** (soft***) | – |
| Tree snag count** (hard***) | |
| Tree snag count** (hard***) |
* Decomposition Classes: N - no or little integration with the soil surface; I - partial to nearly full integration with the soil surface.
** >10.16cm diameter at 1.3716m above ground and >1.8288m height--if less diameter OR height use applicable down wood type; for pinyon and juniper, use 0.3048m above ground.
*** Hard - tree is dead with most or all of bark intact; Soft - most of bark has sloughed off.
Table 8. Canopy structure (% cover)
| Height Above Ground (ft) | Tree | Shrub/Vine | Grass/ Grasslike |
Forb |
|---|---|---|---|---|
| <0.5 | 0% | 0% | 0-3% | 1-3% |
| >0.5 <= 1 | 0% | 0% | 0-25% | 0-15% |
| >1 <= 2 | 0% | 0% | 0-1% | 1-5% |
| >2 <= 4.5 | 0% | 10-20% | 0-1% | 0% |
| >4.5 <= 13 | 0-1% | 5-25% | – | – |
| >13 <= 40 | 0-1% | 0% | – | – |
| >40 <= 80 | 0-5% | – | – | – |
| >80 <= 120 | – | – | – | – |
| >120 | – | – | – | – |
Figure 9. Plant community growth curve (percent production by month). CA1801, Buckbrush (Ceanothus cuneatus). Northern Sierra Nevada Foothills LRU.
| Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | Oct | Nov | Dec |
|---|---|---|---|---|---|---|---|---|---|---|---|
| J | F | M | A | M | J | J | A | S | O | N | D |
| 0 | 5 | 25 | 35 | 20 | 10 | 5 | 0 | 0 | 0 | 0 | 0 |
Community 1.2
Post-fire plant community

Figure 10. Community Phase 1.2, with buckbrush seedlings in post-fire community; Photo: C. Savastio, 2011.
This community phase is characterized by scattered buckbrush (usually <15% canopy cover) that are small and have germinated following scarification from a moderately intense fire (Biswell, 1963; League, 2005). Shrub height is less than three feet. Sites are dominated by annual herbaceous plants, notably the same plants common in community phase 1.1. This community phase is essential in the ecological dynamics of this site. During the first two years following fire, buckbrush seedlings emerge and after three years the shrubs start to gain competitive advantage (League, 2005). Seedling buckbrush roots are able to quickly penetrate much further into the soil profile than the herbaceous plants (League, 2005) even during the first year. Fuel densities for the first ten years would unlikely support another stand replacing fire, but the occurrence of a second fire before the young shrubs start to produce seed would severely decrease the abundance of buckbrush. Fire free periods longer than 30 years would allow sufficient seed bank to accumulate, which has a hard coating and can remain viable for many years (Keeley, 1987) until the occurrence of the next stand replacing fire. Although hotter fires would likely reduce the basal area of foothill pine, some large trees can withstand moderately severe fires (Howard, 1992) because of their self-pruned trunks with thick bark (Lawrence, 1966). More frequent fires would not allow establishing pines to survive, whereas areas that go longer than 100 years without fire might contain higher tree cover. Scattered perennial grasses may establish in the post fire environment or alternatively survive low to medium intensity fires (Simonin, 2001). The small size and sparse leafy material of bottlebrush squirreltail make it a fire-tolerant species; the solid culms of the plant do not burn easily and the growing points below the soil surface often survive (Simonin, 2001). Nevertheless, perennial grasses are minor in biomass compared to the annual forbs and the non-native grasses. Less severe fires might favor the establishment of annual non-native grasses the following spring, as these plants can take advantage of fire related nitrogen in the soil. In some annual grassland systems, the advantage sometimes seen in red brome (Bromus rubens) will be most pronounced three to five years post-fire (Simonin, 2001a; also see B. hordeaceus in Howard, 1998). Keeley (1990) documented cases in which B. rubens altered fire return intervals in chaparral systems, leading to degraded brome dominated plant communities. However, this surge in annual grass production tends to be short-lived in less productive systems and would be expected to be insignificant in this ecological site. The forb component usually includes California goldfields (Lasthenia californica), purple sanicle (Sanicula bipinnatifida), desert parsley (Lomatium spp), bluedics (Dichelostemma capitana), vinegarweed (Trichostema lanceolatum), and annual agoseris (Agoseris heterophylla). Annual grasses often include fescue (Vulpia spp.), soft brome (Bromus hordeaceus), red brome (Bromus rubens), and nit grass (Gastridum pheloides). The main perennial bunchgrass is squirreltail (Elymus elymoides), although trace amounts of melicgrass (Melica spp.), and poa (Poa spp.) can be found in this ecological site.
Figure 11. Annual production by plant type (representative values) or group (midpoint values)
Table 9. Annual production by plant type
| Plant type | Low (lb/acre) |
Representative value (lb/acre) |
High (lb/acre) |
|---|---|---|---|
| Shrub/Vine | 1 | 94 | 182 |
| Forb | 0 | 39 | 125 |
| Grass/Grasslike | 1 | 22 | 56 |
| Total | 2 | 155 | 363 |
Table 10. Soil surface cover
| Tree basal cover | 0% |
|---|---|
| Shrub/vine/liana basal cover | 0-2% |
| Grass/grasslike basal cover | 0-1% |
| Forb basal cover | 0-1% |
| Non-vascular plants | 10-45% |
| Biological crusts | 0% |
| Litter | 13-30% |
| Surface fragments >0.25" and <=3" | 12-35% |
| Surface fragments >3" | 4-20% |
| Bedrock | 1-4% |
| Water | 0% |
| Bare ground | 5-32% |
Table 11. Woody ground cover
| Downed wood, fine-small (<0.40" diameter; 1-hour fuels) | 0-4% |
|---|---|
| Downed wood, fine-medium (0.40-0.99" diameter; 10-hour fuels) | 0-3% |
| Downed wood, fine-large (1.00-2.99" diameter; 100-hour fuels) | 0% |
| Downed wood, coarse-small (3.00-8.99" diameter; 1,000-hour fuels) | 0% |
| Downed wood, coarse-large (>9.00" diameter; 10,000-hour fuels) | 0% |
| Tree snags** (hard***) | – |
| Tree snags** (soft***) | – |
| Tree snag count** (hard***) | |
| Tree snag count** (hard***) |
* Decomposition Classes: N - no or little integration with the soil surface; I - partial to nearly full integration with the soil surface.
** >10.16cm diameter at 1.3716m above ground and >1.8288m height--if less diameter OR height use applicable down wood type; for pinyon and juniper, use 0.3048m above ground.
*** Hard - tree is dead with most or all of bark intact; Soft - most of bark has sloughed off.
Table 12. Canopy structure (% cover)
| Height Above Ground (ft) | Tree | Shrub/Vine | Grass/ Grasslike |
Forb |
|---|---|---|---|---|
| <0.5 | 0% | 0% | 0-1% | 5-20% |
| >0.5 <= 1 | 0% | 0% | 1-27% | 2-10% |
| >1 <= 2 | 0% | 0-8% | 5-20% | 0% |
| >2 <= 4.5 | 0% | 5-43% | 0-2% | 0% |
| >4.5 <= 13 | 0% | 0% | – | – |
| >13 <= 40 | 0-1% | – | – | – |
| >40 <= 80 | – | – | – | – |
| >80 <= 120 | – | – | – | – |
| >120 | – | – | – | – |
Community 1.3
Cleared shrub community

Figure 12. Community Phase 1.3, recently cleared brush (within 3-5 years); Photo: D. Evans, 2013.
This community phase consists of similar herbaceous vegetation as in the surrounding intact shrub communities. The production may depend on several factors, including domestic livestock presence, winter-spring precipitation, and spring temperature, human disturbances (OHV use, mining activity), etc. Nevertheless, a detailed study of both soils and vegetation would be needed to elucidate the ecological dynamics of this community phase, including the time period it would take to return to shrub dominance. It is also unknown how the fire return interval would be affected given the paucity of woody debris (slash piles might increase fire frequency). But, the annual grasses would likely provide sufficient, yet a discontinuous source of fuel for summer or fall burns. Reoccurring intensive grazing (both domestic and wildlife) has the potential to maintain the community in this phase; it may prevent the return of woody vegetation given the pressure on reestablishing shrubs. One concern, for which we currently lack information, is how long can the grassland community dominate without crossing an irreversible threshold? After several years of grazing, buckbrush seed might be lost from the seed bank (see Keeley, 1987). This would make transition to the reference community phase more difficult. Additionally, the effects of deer and other wildlife species on reemerging shrubs might require active restoration necessary, even after the removal of domestic livestock occurred. However, as long as the soil resource base remained intact, this ecological site is probably resilient enough to recover, as seed source from adjacent sites would likely reestablish buckbrush. After buckbrush seeds ripen, they shatter, throwing the seeds as far as 30 feet from the mother plant (Evans et al., 1987). Eventually, once the grazing pressures were removed, the reseeding would probably fill in the gaps created by mechanical clearing. This community phase needs to be reexamined, in order to ascertain that it is indeed another community phase of the historical state, and not an alternate state.
Community 1.4
Mature shrub community

Figure 13. Example of a mature shrub community phase in the Red Hills ACEC., Tuolumne County; Photo: C. Savastio, 2011.
This community phase results from several decades without fire and is composed of mature buckbrush shrubs ranging between 3 and 6 feet in height and occupying 30 to 60% canopy cover and occasional traces of other shrubs such as toyon (Heteromeles arbutifolia) and manzanita (Arctostaphylos spp.). Many of the shrubs have considerable dieback and dead woody material. There may be some younger cohorts of buckbrush, but the majority of the shrub community will be more aged. This community phase also has a greater tree density of the California foothill pine (0-10 canopy cover %). Annual grass production in this community phase may be several times greater than native forb and bunchgrass production. These grasses are often sheltered in the shrub interspaces and add to the flammability of potential fires. Due to the added woody and herbaceous fuel, the main transition pathway consists of high severity fires, which will essential return the community structure to a similar community phase as the cleared shrub (1.3). There is very little chance of low intensity fire, which may result in a patchy mosaic, resulting closer to a post-burn community (1.2). Mechanical removal of some of the detritus may lower the fire hazard. However, slash must either be removed or burned during winter months, which may be a costly process. Rotational grazing of livestock in this community phase may also help restore to the reference community by reducing the cover of annual grasses. Perhaps an integrated strategy of prescribed grazing followed by late season burns could help push this community phase towards the reference condition.
Figure 14. Annual production by plant type (representative values) or group (midpoint values)
Table 13. Annual production by plant type
| Plant type | Low (lb/acre) |
Representative value (lb/acre) |
High (lb/acre) |
|---|---|---|---|
| Shrub/Vine | 30 | 118 | 240 |
| Grass/Grasslike | 10 | 95 | 190 |
| Forb | 1 | 17 | 35 |
| Total | 41 | 230 | 465 |
Table 14. Soil surface cover
| Tree basal cover | 0% |
|---|---|
| Shrub/vine/liana basal cover | 0-2% |
| Grass/grasslike basal cover | 0% |
| Forb basal cover | 0% |
| Non-vascular plants | 0-15% |
| Biological crusts | 0% |
| Litter | 0-60% |
| Surface fragments >0.25" and <=3" | 5-100% |
| Surface fragments >3" | 0-10% |
| Bedrock | 0% |
| Water | 0% |
| Bare ground | 0-25% |
Table 15. Woody ground cover
| Downed wood, fine-small (<0.40" diameter; 1-hour fuels) | 0-15% |
|---|---|
| Downed wood, fine-medium (0.40-0.99" diameter; 10-hour fuels) | 0-5% |
| Downed wood, fine-large (1.00-2.99" diameter; 100-hour fuels) | 0% |
| Downed wood, coarse-small (3.00-8.99" diameter; 1,000-hour fuels) | 0% |
| Downed wood, coarse-large (>9.00" diameter; 10,000-hour fuels) | 0% |
| Tree snags** (hard***) | – |
| Tree snags** (soft***) | – |
| Tree snag count** (hard***) | |
| Tree snag count** (hard***) |
* Decomposition Classes: N - no or little integration with the soil surface; I - partial to nearly full integration with the soil surface.
** >10.16cm diameter at 1.3716m above ground and >1.8288m height--if less diameter OR height use applicable down wood type; for pinyon and juniper, use 0.3048m above ground.
*** Hard - tree is dead with most or all of bark intact; Soft - most of bark has sloughed off.
Table 16. Canopy structure (% cover)
| Height Above Ground (ft) | Tree | Shrub/Vine | Grass/ Grasslike |
Forb |
|---|---|---|---|---|
| <0.5 | 0% | 0% | 0-2% | 0-5% |
| >0.5 <= 1 | 0% | 0% | 5-10% | 1-13% |
| >1 <= 2 | 0% | 0% | 0-7% | 0% |
| >2 <= 4.5 | 0% | 28-35% | 0-20% | 0% |
| >4.5 <= 13 | 0-1% | 30-30% | – | – |
| >13 <= 40 | 0-3% | – | – | – |
| >40 <= 80 | 0-5% | – | – | – |
| >80 <= 120 | – | – | – | – |
| >120 | – | – | – | – |
Pathway 1.1c
Community 1.1 to 1.2
This pathway occurs after patchy, low to moderate severity fire.
Pathway 1.1b
Community 1.1 to 1.3
This pathway occurs after most woody vegetation has been mechanically removed. Alternatively, high severity fires may render a similar change in community structure.
Pathway 1.1a
Community 1.1 to 1.4
This pathway occurs after a considerable amount of time (> 50 years) without fire, under normal growth and development.
Pathway 1.2a
Community 1.2 to 1.1
This pathway occurs over time, pending the absence of fire.
Pathway 1.2b
Community 1.2 to 1.3
Mechanical removal of woody vegetation before the buckbrush have matured may move the community into this phase. This pathway might also occur when intensive grazing occurs early in the post-fire community phase. The combined effects of burning and heavy grazing on buckbrush stands often will stress the establishing shrubs and thus convert shrubland into a grass-forb dominated community (see Biswell, 1963).
Pathway 1.3a
Community 1.3 to 1.2
This pathway occurs with normal time and progression, pending that no major disturbances degrade the soil base. Furthermore, fire occurring in this community phase will greatly enhance buckbrush regeneration, which will hasten the return to community 1.2, the post-fire community.
Pathway 1.1a
Community 1.4 to 1.3
This pathway occurs after a high severity, stand replacing fire. Alternatively, complete brush removal followed by slash burning may cause the community phase changes.
Additional community tables
Table 17. Community 1.1 plant community composition
| Group | Common name | Symbol | Scientific name | Annual production (lb/acre) | Foliar cover (%) | |
|---|---|---|---|---|---|---|
|
Tree
|
||||||
| 1 | Trees | 0–165 | ||||
| California foothill pine | PISA2 | Pinus sabiniana | 0–165 | 0–20 | ||
|
Shrub/Vine
|
||||||
| 2 | Native shrubs | 70–118 | ||||
| buckbrush | CECU | Ceanothus cuneatus | 70–150 | 5–25 | ||
|
Grass/Grasslike
|
||||||
| 3 | Annual grasses | 6–150 | ||||
| soft brome | BRHO2 | Bromus hordeaceus | 2–33 | 1–13 | ||
| fescue | VULPI | Vulpia | 0–3 | 1–10 | ||
| red brome | BRRU2 | Bromus rubens | 0 | 0–1 | ||
| nit grass | GAPH2 | Gastridium phleoides | 0 | 0–1 | ||
| 5 | Native bunchgrasses | 0–5 | ||||
| melicgrass | MELIC | Melica | 0–1 | 0–2 | ||
| squirreltail | ELEL5 | Elymus elymoides | 0 | 0–1 | ||
|
Forb
|
||||||
| 4 | Native forbs | 21–262 | ||||
| goldback fern | PETR7 | Pentagramma triangularis | 0–144 | 0–3 | ||
| bluedicks | DICA14 | Dichelostemma capitatum | 0–76 | 0–13 | ||
| purple sanicle | SABI3 | Sanicula bipinnatifida | 20–40 | 1–8 | ||
| vinegarweed | TRLA4 | Trichostema lanceolatum | 0–24 | 0–2 | ||
| California goldfields | LACA7 | Lasthenia californica | 1–23 | 5–10 | ||
| dotseed plantain | PLER3 | Plantago erecta | 0–18 | 0–5 | ||
| annual agoseris | AGHE2 | Agoseris heterophylla | 0–1 | 0–5 | ||
| bedstraw | GALIU | Galium | 0 | 0–1 | ||
Table 18. Community 1.2 plant community composition
| Group | Common name | Symbol | Scientific name | Annual production (lb/acre) | Foliar cover (%) | |
|---|---|---|---|---|---|---|
|
Shrub/Vine
|
||||||
| 2 | Native shrubs | 1–186 | ||||
| buckbrush | CECU | Ceanothus cuneatus | 1–94 | 5–40 | ||
| toyon | HEAR5 | Heteromeles arbutifolia | 0 | 0–2 | ||
|
Grass/Grasslike
|
||||||
| 3 | Annual grasses | 1–56 | ||||
| red brome | BRRU2 | Bromus rubens | 1–22 | 1–12 | ||
| soft brome | BRHO2 | Bromus hordeaceus | 1–20 | 5–20 | ||
| nit grass | GAPH2 | Gastridium phleoides | 0 | 0–1 | ||
| Pacific fescue | VUMIP | Vulpia microstachys var. pauciflora | 0 | 0–1 | ||
| 5 | Native bunchgrasses | 0–5 | ||||
| squirreltail | ELEL5 | Elymus elymoides | 0–5 | 0–1 | ||
| melicgrass | MELIC | Melica | 1–2 | 1–2 | ||
|
Forb
|
||||||
| 4 | Native forbs | 0–125 | ||||
| bluedicks | DICA14 | Dichelostemma capitatum | 0–36 | 0–14 | ||
| vinegarweed | TRLA4 | Trichostema lanceolatum | 0–20 | 0–6 | ||
| desertparsley | LOMAT | Lomatium | 0–20 | 0–2 | ||
| California goldfields | LACA7 | Lasthenia californica | 0–10 | 0–3 | ||
| purple sanicle | SABI3 | Sanicula bipinnatifida | 0–2 | 2–8 | ||
| annual agoseris | AGHE2 | Agoseris heterophylla | 0–1 | 0–8 | ||
| bedstraw | GALIU | Galium | 0–1 | 0–1 | ||
| trefoil | LOTUS | Lotus | 0–1 | 0–1 | ||
| goldback fern | PETR7 | Pentagramma triangularis | 0–1 | 0–1 | ||
| dotseed plantain | PLER3 | Plantago erecta | 0–1 | 0–1 | ||
Table 19. Community 1.4 plant community composition
| Group | Common name | Symbol | Scientific name | Annual production (lb/acre) | Foliar cover (%) | |
|---|---|---|---|---|---|---|
|
Tree
|
||||||
| 1 | Trees | 0–165 | ||||
| California foothill pine | PISA2 | Pinus sabiniana | 0–165 | 1–5 | ||
|
Shrub/Vine
|
||||||
| 2 | Native shrubs | 30–118 | ||||
| buckbrush | CECU | Ceanothus cuneatus | 30–118 | 28–35 | ||
|
Grass/Grasslike
|
||||||
| 3 | Annual grasses | 0–95 | ||||
| nit grass | GAPH2 | Gastridium phleoides | 0–73 | 1–5 | ||
| red brome | BRRU2 | Bromus rubens | 0–20 | 1–10 | ||
| wild oat | AVFA | Avena fatua | 0 | 1–20 | ||
| soft brome | BRHO2 | Bromus hordeaceus | 0 | 1–7 | ||
| 5 | Native bunchgrasses | 0–5 | ||||
| squirreltail | ELEL5 | Elymus elymoides | 0–1 | 0–1 | ||
| melicgrass | MELIC | Melica | 0–1 | 0–1 | ||
|
Forb
|
||||||
| 4 | Native forbs | 0–35 | ||||
| vinegarweed | TRLA4 | Trichostema lanceolatum | 0–16 | 0 | ||
| Indian paintbrush | CASTI2 | Castilleja | 0 | 1–10 | ||
| bedstraw | GALIU | Galium | 0 | 0–1 | ||
| goldback fern | PETR7 | Pentagramma triangularis | 0 | 0–1 | ||
| dotseed plantain | PLER3 | Plantago erecta | 0 | 0 | ||
| poison sanicle | SABI2 | Sanicula bipinnata | 0 | 0 | ||
| California goldfields | LACA7 | Lasthenia californica | 0 | 0 | ||
| tarweed | MADIA | Madia | 0 | 0 | ||
| soapplant | CHLOR3 | Chlorogalum | 0 | 0 | ||
Interpretations
Animal community
Various wildlife species have been observed in this site. The Red Hills is a winter roosting site of the bald eagle (Haliaeetus leucocephalus). At least two of the streams support aquatic life such as the Red Hills roach (Lavinia symmetricus) and mosquito fish (Gambusia affinis) (US Department of Interior, 2013). This area has close to 100 bird species during some time of the year. Common mammals include bobcat (Felis rufus), coyote, (Canis latrans), Western gray squirrel (Spermophilus beecheyi), and mule deer (Odocoileus hemionus); reptiles such as Western pond turtle (Clemnys marmorata), Sierra garter snake (Thamnophis couchi couchi)
California king snake (Lampropeltis getulus californiae), and Northern Pacific rattlesnake (Crotalus virdis oreganus) have been spotted.
Hydrological functions
The soils ecological site functions largely as subsurface recharge although much of the moisture may be shed as surface runoff during higher precipitation events as a result of relatively low water permeability of the surface. Erosive rainfall events are not uncommon during the fall with flow patterns being very common, terracettes being somewhat common, and patches of effective bare ground being many, small and generally interconnected.
Moisture may discharge over time to lower points of this landscape moving along bedrock features and through bedrock fractures and nearing or emerging from the soil surface along slope breaks where restrictive subsurface parent material redirects subsurface flow laterally rather than vertically. The amount of runoff may be slightly lower than in the adjacent blue oak grasslands in the area. Soil loss would be expected to be similar to adjacent ecological sites in MLRA 18. The exception being in areas of recent burn, where lack of cover and more exposed soil will contribute to greater loss. This would also be true of areas in the blue oak woodland that experience burns and loose surface litter and plant cover. The mature shrub plant community phase has lower sediment yield and soil loss than the other community phases. Therefore, a mixture of community phases on the landscape may ensure a functional ecosystem.
Recreational uses
This site is often used in recreation, such as horseback riding, bird viewing, recreational botany, and hiking.
Wood products
Little to none. Foothill pine may be used as fire wood.
Supporting information
Type locality
| Location 1: Tuolumne County, CA | |
|---|---|
| Township/Range/Section | T1S R14E S34 |
| UTM zone | N |
| UTM northing | 4193107 |
| UTM easting | 722167 |
| Latitude | 37° 51′ 30″ |
| Longitude | 120° 28′ 29″ |
| General legal description | About ½ mile south of Chinese Camp, California. |
Other references
Alexander, E. B. and J. Dushey. 2011. Topographic and soil differences from peridotite to serpentinite. Geomorphology 135:271-276.
Biswell, H. H. 1963. Research in wildland fire ecology in California. In: Proceedings, 2nd annual Tall Timbers fire ecology conference; 1963 March 14-15; Tallahassee, FL. No. 2. Tallahassee, FL: Tall Timbers Research Station: 63-97.
Bolsinger, C. L. 1988. The hardwoods of California's timberlands, woodlands, and savannas. Res. Bull. PNW-RB-148. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station. 148 p
Brooks, R. 1987. Serpentine and its vegetation. Discorides Press: Portland, OR.
Cleland, D.T., Avers, P.E., McNab, W.H., Jensen, M.E., Bailey, R.G., and W.E. Russell. 1997. National hierarchical framework of ecological units. In: M.S. Boyce and A. Hanley, eds. Ecosystem Management: Applications for Sustainable Forest and Wildlife Resources. New Haven, CT. Yale University Press. Pp 181-200.
Delwiche, C. C., P. J. Zinke, and C. M. Johnson. 1965. Nitrogen fixation by ceanothus. Plant Pathology 40:1045-1047.
Evans, R. A., H. H. Biswell, and D. E. Palmquist. 1987. Seed dispersal in Ceanothus cuneatus and C. leucodermis in a Sierran oak-woodland savanna. Madroño 34 (4): 282-293.
Gram, W., E. Borer, K. Cottingham, E. Seabloom, V. Bloucher, L. Goldwasser, F. Micheli, B. Kendall, and R. Burton. 2004. Distribution of plants in a California serpentine grassland: are rocky hummocks spatial refuges for native species? Plant Ecology 172:159-171.
Harrison, S., B. Inouye, and H. Safford. 2003. Ecological heterogeneity in the effects of grazing and fire on grassland diverstity. Conservation Biology 17:837-845.
Hobbs, R. J., S. Yates, and H. A. Mooney. 2007. Long-term data reveal complex dynamics in grassland in relation to climate and disturbance. Ecological Monographs 77(4):545-568.
Howard, J. L. 1992. Pinus sabiniana. In: Fire Effects Information System, [Online].
U.S. Department of Agriculture, Forest Service,
Rocky Mountain Research Station, Fire Sciences Laboratory (Producer).
Available: http://www.fs.fed.us/database/feis/ [2013, March 15].
Howard, J. L. 1998. Bromus hordeaceus. In: Fire Effects Information System, [Online].
U.S. Department of Agriculture, Forest Service,
Rocky Mountain Research Station, Fire Sciences Laboratory (Producer).
Available: http://www.fs.fed.us/database/feis/ [2013, April 9].
Huenneke, L., S. Hamburg, R. Koide, H. Mooney, and P. Vitousek. 1990. Effects of soil resources on plant invasion and community structure in Californian serpentine grassland ecology. Ecology 71:478-491.
Hufford, K. M., S. J. Mazer, and M. D. Camara. 2008. Local adaptation and effects of grazing among seedlings of two native California bunchgrass species: Implications for restoration. Restoration Ecology 16:56-69.
Hutchings, S. S. and G. Stewart. 1953. Increasing forage yields and sheep production on Intermountain winter ranges. Circular No. 925. Washington, DC: U.S. Department of Agriculture. 63 p.
Jackson, L. 1985. Ecological origins of California’s Mediterranean grasses. Journal of Biogeography 12:349-361.
Keeley, J. E. 1987. Role of fire in seed germination of woody taxa in California chaparral. Ecology 68(2): 434-443.
Keeley, J. E. 1990. The California valley grassland In: Schoenherr, A. Allan (ed.). Endangered plant communities of southern California: Proceedings of the 15th annual symposium; 1989 October 28, Fullerton, CA, Special Publication No. 3: Claremont, CA: Southern California Botanists 2-23.
Kruckeberg, A. R. 1984. California serpentines: flora, vegetation, geology, soils, and management problems. University of California Press: Berkeley and Los Angeles, CA.
Lawrence, G. E. 1966. Ecology of vertebrate animals in relation to chaparral fire in the Sierra Nevada foothills. Ecology 47(2): 278-291.
Lazarus, B. E., J. H. Richards, V. P. Claassen, R. E. O’Dell, and M. A. Ferrell. 2011. Species and specific plant-soil interactions influence plant distribution on serpentine soils. Plant and Soil 342:327-344.
League, K. R. 2005. Ceanothus cuneatus. In: Fire Effects Information System, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer). Available: http://www.fs.fed.us/database/feis/ [2013, March 15].
McClaran, M.P. 1986. Age structure of Quercus douglasii in relation to livestock grazing and fire. Ph.D. Dissertation. Univ. of Calif., Berkeley. 119 pp.
McGahan, D. G., R. J. Southard, and V. P. Claassen. 2009. Plant-Available calcium varies widely in soils on serpentinite landscapes. Soil Science Society of America Journal 73:2087-2095.
McNab, W.H., Cleland, D.T., Freeouf, J.A., Keys, Jr., Nowacki, G.J., and C. A. Carpenter. 2007. Descriptions of ecological subregions: sections of the coterminous United States. GTR-WO-76B. [CD-ROM] Washington, DC. US. Dept. Agric., For. Serv. 80 pg.
Omernick, J. M., and G.E. Griffith. 2007. Ecoregions of the Coterminous United States: Evolution of a Hierarchical Spatial Framework.
Parsons, D. J., P. W. Rundel, R. P. Hedlund, and G. A. Baker. 1981. Survival of severe drought by a non-sprouting chaparral shrub. American Journal of Botany 68(7):973-979.
Regan H. M., J. B. Crookston, R. Swab, J. Franklin, and D. M. Lawson. 2010. Habitat fragmentation and altered fire regime create trade-offs for an obligate seedling shrub. Ecology 91(4):1114-1123.
Safford, H. D. and S. Harrison. 2004. Fire effects on plant diversity in serpentine vs. sandstone chaparral. Ecology 85:539-548.
Simonin, K. A. 2001. Elymus elymoides. In: Fire Effects Information System, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer). Available: http://www.fs.fed.us/database/feis/ [2014, March 12].
Simonin, K. A. 2001a. Bromus rubens, Bromus madritensis. Simonin, Kevin A. 2001. Bromus rubens, Bromus madritensis. In: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer). Available: http://www.fs.fed.us/database/feis/ [2014, April 29].
Stewart, O. C., H. T. Lewis (ed.) and M. K. Anderson (ed.) 2002. Forgotten fires: Native Americans and the transient wilderness. University of Oklahoma Press: Norman, OK.
US Department of the Interior Bureau of Land Management. 2013. The Red Hills of Tuolumne County. Available: http://www.blm.gov/ca/st/en/fo/folsom/redhills/redhillshomepg1.html
[2013, April 1]
Woodruff, L. G., W. F. Cannon, D. D. Eberl, D. B. Smith, J. E. Kilburn, J. D. Horton, R. G. Horton, R. G. Garrett, and R. A. Klassen. 2009. Continental-scale patterns in soil geochemistry and mineralogy: results from two transects across the United States and Canada. Applied Geochemistry 24:1369-1381.
Contributors
Dave Evans
Rangeland health reference sheet
Interpreting Indicators of Rangeland Health is a qualitative assessment protocol used to determine ecosystem condition based on benchmark characteristics described in the Reference Sheet. A suite of 17 (or more) indicators are typically considered in an assessment. The ecological site(s) representative of an assessment location must be known prior to applying the protocol and must be verified based on soils and climate. Current plant community cannot be used to identify the ecological site.
| Author(s)/participant(s) | |
|---|---|
| Contact for lead author | |
| Date | |
| Approved by | |
| Approval date | |
| Composition (Indicators 10 and 12) based on | Annual Production |
Indicators
-
Number and extent of rills:
-
Presence of water flow patterns:
-
Number and height of erosional pedestals or terracettes:
-
Bare ground from Ecological Site Description or other studies (rock, litter, lichen, moss, plant canopy are not bare ground):
-
Number of gullies and erosion associated with gullies:
-
Extent of wind scoured, blowouts and/or depositional areas:
-
Amount of litter movement (describe size and distance expected to travel):
-
Soil surface (top few mm) resistance to erosion (stability values are averages - most sites will show a range of values):
-
Soil surface structure and SOM content (include type of structure and A-horizon color and thickness):
-
Effect of community phase composition (relative proportion of different functional groups) and spatial distribution on infiltration and runoff:
-
Presence and thickness of compaction layer (usually none; describe soil profile features which may be mistaken for compaction on this site):
-
Functional/Structural Groups (list in order of descending dominance by above-ground annual-production or live foliar cover using symbols: >>, >, = to indicate much greater than, greater than, and equal to):
Dominant:
Sub-dominant:
Other:
Additional:
-
Amount of plant mortality and decadence (include which functional groups are expected to show mortality or decadence):
-
Average percent litter cover (%) and depth ( in):
-
Expected annual annual-production (this is TOTAL above-ground annual-production, not just forage annual-production):
-
Potential invasive (including noxious) species (native and non-native). List species which BOTH characterize degraded states and have the potential to become a dominant or co-dominant species on the ecological site if their future establishment and growth is not actively controlled by management interventions. Species that become dominant for only one to several years (e.g., short-term response to drought or wildfire) are not invasive plants. Note that unlike other indicators, we are describing what is NOT expected in the reference state for the ecological site:
-
Perennial plant reproductive capability:
Print Options
Sections
Font
Other
The Ecosystem Dynamics Interpretive Tool is an information system framework developed by the USDA-ARS Jornada Experimental Range, USDA Natural Resources Conservation Service, and New Mexico State University.
Click on box and path labels to scroll to the respective text.