
Natural Resources
Conservation Service
Ecological site F022AE025CA
Loamy Moist Outwash
Accessed: 11/21/2024
General information
Approved. An approved ecological site description has undergone quality control and quality assurance review. It contains a working state and transition model, enough information to identify the ecological site, and full documentation for all ecosystem states contained in the state and transition model.
Figure 1. Mapped extent
Areas shown in blue indicate the maximum mapped extent of this ecological site. Other ecological sites likely occur within the highlighted areas. It is also possible for this ecological site to occur outside of highlighted areas if detailed soil survey has not been completed or recently updated.
MLRA notes
Major Land Resource Area (MLRA): 022A–Sierra Nevada and Tehachapi Mountains
Major Land Resource Area 22A, Sierra Nevada Mountains, is located predominantly in California and a small section of western Nevada. The area lies completely within the Sierra Nevada Section of the Cascade-Sierra Mountains Province. The Sierra Nevada range has a gentle western slope, and a very abrupt eastern slope. The Sierra Nevada consists of hilly to steep mountains and occasional flatter mountain valleys. Elevation ranges between 1,500 and 9,000 ft throughout most of the range, but peaks often exceed 12,000 ft. The highest point in the continental US occurs in this MLRA (Mount Whitney, 14,494 ft). Most of the Sierra Nevada is dominated by granitic rock of the Mesozoic age, known as the Sierra Nevada Batholith. The northern half is flanked on the west by a metamorphic belt, which consists of highly metamorphosed sedimentary and volcanic rocks. Additionally, glacial activity of the Pleistocene has played a major role in shaping Sierra Nevada features, including cirques, aretes, and glacial deposits and moraines. Average annual precipitation ranges from 20 to 80 inches in most of the area, with increases along elevational and south-north gradients. The soil temperature regime ranges from mesic, frigid, and cryic.
LRU "E" Northern Sierran Upper Montane: This LRU occurs at the mid elevations of the Sierra Nevada, from the Sonora Pass area to the higher mountains in the vicinity of Quincy. Elevations are typically between 5,500 feet to 8,500 feet, with the lower elevations typically on southern aspects, and the higher elevations on northern aspects. The frost-free season is 60 to 125 days, MAAT ranges from 40 to 50 F, and MAP ranges from 35 to 85 inches. The soil temperature regime is mostly frigid, with some cryic soil temperatures at the upper elevations and northern aspects. Soil moisture regimes are mostly xeric, but may be udic where snow persists through spring.
Ecological site concept
This site occurs on valley bottoms on gently sloping glacial outwash, alluvial fans and old lake terraces with deep to very deep soils formed in colluvium and alluvium from mixed sources. Soils have loamy textures and relatively high soil moisture holding capacity and nutrients. Elevations range from 6220 to 6710 feet and slopes typically between 0 and 9 percent. Jeffrey pine (Pinus jeffreyi) dominates a productive mixed-conifer forest, with white fir (Abies concolor), sugar pine (Pinus lambertiana), and incense cedar (Calocedrus decurrens) important secondary canopy species. Thimbleberry (Rubus parviflorus) and creeping snowberry (Symphoricarpos mollis) are the dominant shrub species. Thimbleberry prefers moist, fertile soils and is indicative of the additional soil moisture in the loamy soils of this ecological site. This additional moisture allows for greater understory diversity than similar topographic positions with sandy soils, and more than 40 species of forbs occur on this site.
Associated sites
| F022AC006CA |
Moderately Deep Cryic Sandy Till Occurs on adjacent south-facing cryic mountain slopes with moderately deep loamy-skeletal soils. The vegetation is an open lodgepole pine (Pinus contorta var. murrayana) forest with red fir (Abies magnifica) and western white pine (Pinus monticola). |
|---|---|
| F022AE007CA |
Frigid, Sandy, Moraines And Hill Slopes Occurs on adjacent moraines and hillslopes with sandy, moderately deep to very deep soils. The vegetation is white fir (Abies concolor) - Jeffrey pine (Pinus jeffreyi) forest. |
| F022AE013CA |
Frigid, Loamy, Volcanic Mountain Slopes Occurs on adjacent mountain slopes with moderately deep to deep soils derived from andesite. The vegetation is a white fir (Abies concolor) - mixed conifer forest. Jeffrey pine (Pinus jeffreyi), sugar pine (Pinus lambertiana) and incense cedar (Calocedrus decurrens) are all important species. |
| R022AX107CA |
Frigid C Channel System This riparian complex occurs along C-B type channels with gravelly to cobbly channel substrates and 2 - 3 percent slopes. The vegetation is characterized by willow (Salix ssp.) - aspen (Populus tremuloides) - cottonwood (Populus balsamifera) communities. |
Similar sites
| F022AF002CA |
Frigid, Sandy, Or Loamy Outwash Occurs in the "AF" lru where precipitation is lower. Jeffrey pine (Pinus jeffreyi) dominates the forest canopy and greenleaf manzanita (Arctostaphylos patula) and prostrate ceanothus (Ceanothus prostratus) are dominant understory species. |
|---|---|
| F022AF006CA |
Loamy Frigid Metamorphic Slopes Occurs in the "AF" lru, where precipitation is lower. Forest canopy composition is less diverse, and is strongly dominated by Jeffrey pine (Pinus jeffreyi). |
| F022AE008CA |
Frigid Loamy Moraine Slopes Occurs on similar landforms where cooler and moister topographic positions allow red fir (Abies magnifica) to co-dominate with white fir (Abies concolor). Jeffrey pine (Pinus jeffreyi) is a minor species if present. |
| F022AE013CA |
Frigid, Loamy, Volcanic Mountain Slopes Occurs on mountain slopes. Forest canopy composition is similar, but the understory is less diverse. Greenleaf manzanita (Arctostaphylos patula) and bush chinquapin (Chrysolepis sempervirens) are the dominant understory species. |
| F022AE007CA |
Frigid, Sandy, Moraines And Hill Slopes Occurs on similar landforms and topographic positions with sandy soils. The forest canopy composition is similar, but the understory is less diverse due to lower soil moisture holding capacity of sandy soils. |
Table 1. Dominant plant species
| Tree |
(1) Pinus jeffreyi |
|---|---|
| Shrub |
(1) Rubus parviflorus |
| Herbaceous |
Not specified |
Physiographic features
This ecological site is on gently sloping alluvial fans, lake terraces and moraines. Slopes may range from 0 to 15 percent, but are typically below 9 percent. It is found on all aspects and elevations range from 6220 to 6710 feet. Runoff class is low to medium. This ecological site occurs primarily on the western shores of the Lake Tahoe basin, but is occasionally found on moist loamy soils on the east shore.
Table 2. Representative physiographic features
| Landforms |
(1)
Moraine
(2) Alluvial fan (3) Lake terrace |
|---|---|
| Flooding frequency | None |
| Ponding frequency | None |
| Elevation | 1,896 – 2,045 m |
| Slope | 0 – 15% |
Climatic features
The average annual precipitation ranges from 23 to 43 inches, mostly in the form of snow in the winter months (November through April). The average annual air temperature ranges from 39 to 46 degrees Fahrenheit. The frost-free (>32F) season is 30 to 90 days, and the freeze-free (>28F) season is 80 to 140 days.
Table 3. Representative climatic features
| Frost-free period (average) | 110 days |
|---|---|
| Freeze-free period (average) | 60 days |
| Precipitation total (average) | 838 mm |
Figure 2. Monthly precipitation range
Figure 3. Monthly average minimum and maximum temperature
Figure 4. Annual precipitation pattern
Figure 5. Annual average temperature pattern
Influencing water features
This ecological site is not influenced by wetland or riparian water features.
Soil features
The soils associated with this ecological site are deep to very deep, and formed in colluvium over till derived from mixed parent materials, or are derived from andesite over lacustrine deposits. They are moderately to well drained with very slow to moderately rapid permeability. The soil moisture regime is typic xeric and the soil temperature regime is frigid. Surface rock fragments smaller than 3 inches in diameter average 1 percent cover, and larger fragments also average 1 percent. Surface textures are stony sandy loam and gravelly coarse sandy loam. A thin layer of partially decomposed litter (Oi) overlies the mineral subsurface horizons. Subsurface textures are gravelly, extremely cobbly, and very gravelly coarse sandy loam, stony sandy loam, loam, sandy clay loam, and clay loam. Subsurface rock fragments smaller than 3 inches in diameter range from 0 to 35 percent by volume, and larger fragments range from 0 to 50 percent (for a depth of 0 to 66 inches). The soils correlated to this site include Kingsbeach (fine-loamy, isotic, frigid Ultic Palexeralfs), and Tallac (loamy-skeletal, mixed, superactive, frigid Humic Dystroxerepts). Kingsbeach soils are very deep and derived from andesite over lacustrine deposits. Tallac (Moderately well drained phase) soils are deep to very deep to a duripan, and formed from mixed parent material weathered from glacial deposits.
This ecological site has been correlated with the following mapunits and soil components in the Tahoe Basin soil survey area (CA693):
Musym ; MUname ; Compname ; Local_phase ; Comp_pct
7161 ; Kingsbeach stony sandy loam, 2 to 15 percent slopes ; Kingsbeach ; ; 80
7524 ; Tallac gravelly coarse sandy loam, moderately well drained, 0 to 5 percent slopes ; Tallac ; moderately well drained ; 80; Tallac ; moderately well drained ; 10
7525 ; Tallac gravelly coarse sandy loam, moderately well drained, 5 to 9 percent slopes ; Tallac ; moderately well drained ; 80; ; Tallac ; moderately well drained ; 10
7526 ; Tallac gravelly coarse sandy loam, moderately well drained, 2 to 9 percent slopes, rubbly ; Tallac ; moderately well drained ; 10
7141 ; Inville gravelly coarse sandy loam, 2 to 9 percent slopes, stony ; Kingsbeach ; ; 2
7521 ; Tallac gravelly coarse sandy loam, 5 to 15 percent slopes, very stony ; Tallac ; moderately well drained ; 9
Table 4. Representative soil features
| Surface texture |
(1) Stony sandy loam (2) Gravelly coarse sandy loam |
|---|---|
| Family particle size |
(1) Loamy |
| Drainage class | Moderately well drained to well drained |
| Permeability class | Very slow to moderately rapid |
| Soil depth | 99 cm |
| Surface fragment cover <=3" | 1% |
| Surface fragment cover >3" | 1% |
| Available water capacity (0-101.6cm) |
6.1 – 18.54 cm |
| Soil reaction (1:1 water) (0-101.6cm) |
5.1 – 6.5 |
| Subsurface fragment volume <=3" (Depth not specified) |
0 – 35% |
| Subsurface fragment volume >3" (Depth not specified) |
0 – 50% |
Ecological dynamics
Abiotic Factors
This site occurs on valley bottoms on gently sloping glacial outwash, alluvial fans and old lake terraces with deep to very deep soils formed in colluvium and alluvium from mixed sources. Soils have loamy textures and relatively high soil moisture holding capacity and nutrients. Elevations range from 6220 to 6710 feet and slopes range from 0 to 15 percent. Jeffrey pine dominates a productive mixed-conifer forest, with white fir, sugar pine, and incense cedar important secondary canopy species. Thimbleberry and creeping snowberry are the dominant shrub species. Thimbleberry prefers moist habitats with nutrient-rich soils (Gucker 2012), and is indicative of additional soil moisture in the loamy soils of this ecological site. This additional moisture allows for greater understory diversity than similar topographic positions with sandy soils, and more than 40 species of forbs occur on this site.
Ecological factors
Fire and fire suppression, logging, drought and pathogens are the primary disturbance factors affecting the dynamics of this ecological site. Pre-European settlement, the most successionally advanced community phase was most likely dominated by large old growth Jeffrey pine, with old growth white fire, sugar pine, and incense cedar sharing the canopy. The canopy was relatively open, allowing for a high diversity of shrubs and forbs in the understory (e.g. Beardsley et al. 1999, Murphy and Knopp 2000, Barbour et al. 2002, Taylor 2004, Stephens and Fry 2005, Binkley et al. 2007). Historically, this community phase developed with a mixed fire severity regime, with patchy, frequent, low to medium intensity surface fires, and occasional larger high severity fires, all of which occurred primarily in the fall when fuel moisture was lowest and trees were dormant (Beaty and Taylor 2008). Fire scar analysis indicates the median historic fire return interval for low and moderate severity fires in mixed conifer forests in the Lake Tahoe area was 3 to 9 years, and that larger fires occurred every 12 to 34 years (Beaty and Taylor 2008). This mixed severity fire regime created a forest with a mosaic of age-classes, and provided frequent recruitment opportunities for Jeffrey pine and sugar pine, which require open sites for seedling establishment. Frequent fires kept the understory largely clear of shade-tolerant but fire-intolerant white fir, red fir, and incense cedar, but longer intervals between fires allowed these species to reach sizes where they are less susceptible to fire-induced mortality.
The old-growth phase is currently rare due to either fire suppression or clear-cutting. This ecological site was almost entirely clear-cut during the 1870s to 1890s during the period known as the Comstock Era (Elliot-Fisk et al. 1996, Murphy and Knopp 2000, Barbour et al. 2002, Taylor 2004, Beaty and Taylor 2008). Young forests that have subsequently developed have higher density and basal area, and are comprised of younger and smaller trees with a more even age-class distribution, with most canopy trees 80 to 120 years old (Taylor 2004, Stephens and Fry 2005, Beaty and Taylor 2008). A long-term policy of fire suppression has impacted both these second-growth forests, and the few remaining contemporary stands of old-growth forest (Barbour et al. 2002, Stephens and Fry 2005, Beaty and Taylor 2008). White and red fir and incense cedar have increased in the understory, leading to densely stocked forests with increasing canopy closure, and a build-up of coarse woody debris. The lack of canopy openings limits recruitment opportunities for Jeffrey pine and sugar pine, which decline in importance in these infilled forests. Increasing canopy cover, and lack of bare ground and nutrient cycling also reduces the abundance and richness of the understory in forests with a long duration of fire suppression (e.g. Huisinga et al. 2005, Laughlin et al. 2005, Binkley et al. 2007). Understory trees provide ladder fuels, and the accumulation of highly flammable downed wood increases the likelihood of large high severity canopy fire, and reduces the likelihood that the natural fire regime, which includes frequent low to moderate severity, small to medium-sized fires, can occur. However, management practices such as thinning with prescribed fire can mimic natural processes and help restore these forests back to a more natural condition.
Contemporary forests, with more crowded conditions, and a higher frequency of drought (e.g. Jones et al. 2004) are more susceptible to pathogen induced mortality (Barbour et al. 2002). Bark beetles (Dedroctonus spp.) are significant disease agents for Jeffrey pine and sugar pine. Fire damage increases the likelihood of bark beetle infestation and mortality (Bradley and Tueller 2001, Maloney et al. 2008, Fettig et al. 2010). Drought also increases the likelihood of mortality. Barbour et al. (2002) found that most of the mortality of old-growth Jeffrey pine in the Lake Tahoe Basin was due to severe drought from 1988-1992, and all dead trees were infected by bark beetle. Nitrogen deposition and ozone pollution have been shown to contribute to Jeffrey pine susceptibility to pathogens and mortality in Southern California (e.g. Peterson et al. 1987), but equivalent studies have not been done in the northern Sierra. White pine blister rust (Cronartium ribicola) is a serious threat to sugar pine (Pinus lambertiana) (Murphy and Knopp 2000). White pine blister rust kills old, young, and seedling trees. It can severely inhibit regeneration in infested areas by greatly reducing the sugar pine population. In this ecological site where sugar pine is already present at low abundances, white pine blister rust could eliminate sugar pine altogether. These pathogens may also have played a role in diminishing the importance of pines relative to fir species in contemporary forests (Beardsley et al. 1999).
The reference state consists of the pre-settlement, most successionally advanced community phase (numbered 1.1), and the community phases that result from natural and human disturbances. Community phase 1.1 is deemed the phase representative of the most successionally advanced pre-European plant/animal community including periodic natural surface fires that influenced its composition and production. Because this phase is determined from reconstruction and/or historic literature, some speculation is necessarily involved in describing it.
All tabular data listed for a specific community phase within this ecological site description represent a summary of one or more field data collection plots taken in modal communities within the community phase. Although such data are valuable in understanding the phase (kinds and amounts of ground and surface materials, canopy characteristics, community phase overstory and understory species, production and composition, and growth), they do not represent the absolute range of characteristics or an exhaustive listing of all species that may occur in that phase over the geographic range of the ecological site.
State and transition model
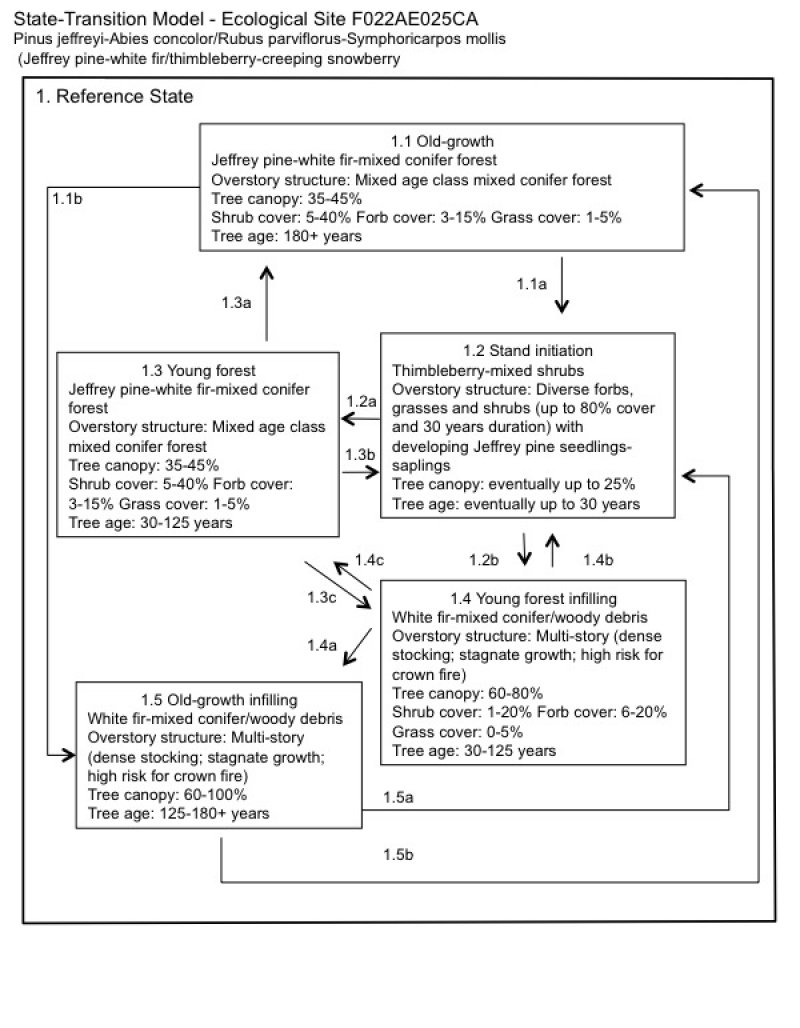
Figure 6. F022AE025CA
More interactive model formats are also available.
View Interactive Models
More interactive model formats are also available.
View Interactive Models
Click on state and transition labels to scroll to the respective text
Ecosystem states
State 1 submodel, plant communities
Communities 1, 5 and 2 (additional pathways)
State 1
Reference
Community 1.1
Old-growth forest
The most successionally advanced community phase is characterized by a multi-aged forest dominated by old-growth Jeffrey pine, with white fir, sugar pine, and incense cedar important secondary species. Red fir is present at higher elevations. The structure of these old growth mixed conifer forests was complex, with both scattered or clumped large (>30 inch diameter) and tall (>120 feet) trees. Among the canopy trees were several understory layers of trees. White fir and incense cedar were most common in the understory and mid-canopy. Understory shrub and forb richness was high and concentrated in canopy openings, with thimbleberry and creeping snowberry the most abundant shrub species.
Community 1.2
Stand initiation
A productive and diverse shrub and forb community that thrives in the openings created by large fires that burn the forest canopy characterizes the stand initiation phase. Although most canopy trees are killed by canopy fire, occasional remnant mature trees may remain. Annual and perennial forbs and perennial grasses dominate the first one to three years after severe fire. Annual forbs including Torrey’s blue-eyed Mary (Collinsia torreyi) and spreading gunsmoke (Gayophytum diffusum) may be productive and abundant in bare, open patches in the first several years following fire (Wright 1985, Schoennagel et al. 2004, Dhaemers 2006, Wayman and North 2007). Silvery lupine (Lupinus argenteus), rose thistle (Cirsium andersonii), and mountain monardella (Monardella odoratissima) colonize from seed. Silvery lupine is a nitrogen fixer, and an important colonizer of disturbed sites, and is abundant when fire return intervals are short (Schoennagel et al. 2004). This species increases in vigor relative to pre-burn status by the second year after fire (Rau et al. 2008). Perennials that are only top-killed by fire and that spread by rootstocks such as Browns peony (Paeonia brownii), lambstongue ragwort (Senecio intigerrimus), woolly mules-ears (Wyethia mollis), arrowleaf balsamroot (Balsamorhiza sagittata), and dusky onion (Allium campanulatum) will increase in abundance by the second or third season after fire. Woolly mules-ears can greatly increase after fire, sometimes becoming dominant (Parker and Yoder-Williams 1989, Riegel et al. 2002). Squirreltail (Elymus elymoides) is top killed by fire and will resprout from the root crown (Simonin 2001). Ross sedge (Carex rossii) is a colonizer after fire, and can regenerate from surviving rhizomes or from heat-activated seed stored in the soil (Anderson 2008). Shrubs will begin to resprout and germinate from seed as early as the first year post-fire, gaining dominance with time. Thimbleberry resprouts from rhizomes or from long-lived seed stored in the soil seedbank (Gucker 2012). Thimbleberry sprouts or seedlings will be present as soon as the first year after fire or clearcut, peaking in abundance 8 to 11 years after disturbance (Gucker 2012). Although thimbleberry abundance does decline with increasing canopy closure, it will remain relatively abundant in canopy openings in this site, and can persist in closed canopy forest (Gucker 2012). Creeping snowberry is capable of resprouting from rhizomes after disturbance (Snyder 1991), and there are reports of increased abundance after prescribed fire (Zhang et al. 2008), and after forest thinning (Lindh and Muir 2004); however, it will likely be less important in this community phase relative to more aggressive disturbance responders such as thimbleberry. Fire dependent shrubs such as greenleaf manzanita (Arctostaphylos patula), Saskatoon serviceberry (Amelanchier alnfiolia), Utah serviceberry (A. utahensis), and mountain whitethorn (Ceanothus cordulatus) resprout and/or germinate from seed vigorously after a fire. Greenleaf manzanita resprouts from underground lignotubers, and regenerates from heat scarified seeds that may survive in the soil for more than 400 years (Nagal and Taylor 2005, Hauser 2007). Mountain whitethorn is an obligate resprouter after low to medium intensity fire, and seeds require heat for germination (Reeves 2006). Saskatoon serviceberry may resprout from the root crown or rhizomes after low to moderate severity fire, and from deeply buried rhizomes after high severity fire (Fryer 1997). Scoulers willow (Salix scouleriana) is a disturbance-adapted species that will resprout from the root crown after fire or mechanical disturbance, and colonize from off-site seed sources, being especially successful where fire or disturbance has exposed mineral soil (Anderson 2001). Fire intolerant shrubs such as antelope bitterbrush (Purshia tridentata) and mountain big sagebrush (Artemisia tridentata ssp. vaseyana) will be killed by fire, and are generally not present or are very minor species in the stand initiation phase. Jeffrey pine and sugar pine require bare soil and an open canopy to regenerate, and seedlings will sprout following fire, but may take 30 years to begin to dominate over the shrubland community phase (Smith 1994, Azuma et al. 2004). Incense cedar can establish in bare mineral soil with partial shade or in full shade under heavy tree or shrub cover (Tollefson 2008). White fir seedlings are most successful with full shade (Zouhar 2001), and begin to establish once shrub and other tree cover is well established.
Community 1.3
Young forest

Figure 7. Community Phase 1.3
This young open mixed conifer forest community phase develops with the natural fire regime, or with manual thinning and prescribed fire treatment. Low to moderate intensity fire clears the understory and removes fuel before it reaches hazardous levels, although severe high intensity canopy fires are also possible. The young forest community phases (1.3 and 1.4) are the most common expressions of this ecological site at this time in the Lake Tahoe Basin. The young, open mixed conifer community phase is a heavily managed forest. Manual thinning and prescribed burns reduce the white fir component, reduce fuel loads, and create canopy openings in the forest. Natural fires are generally quickly extinguished in this forest because of its proximity to urban areas.
Forest overstory. Jeffrey pine and white fir are co-dominant in the overstory, and incense cedar and sugar pine are important secondary canopy species. Patches of quaking aspen (Populus tremuloides) may be present. All four conifer species are also present in the mid-canopy layer, with white fir and incense cedar most abundant in this layer. This community phase begins with 10 to 20-foot tall pole-sized trees and matures to 120-foot tall tress with diameters ranging from 19 to 30 inches. Canopy cover ranges from 35 to 45 percent, with an average of 40 percent cover.
Forest understory. An open forest canopy, and frequent low to moderate severity fires allow for a relatively high cover and high diversity of shrubs and forbs in the understory of this community. Shrub cover averages 15 percent, and ranges from 5 to 40 percent. Forb cover averages 10 percent, and ranges from 3 to 14 percent. Grasses and grasslike species make a minor contribution to the understory, with an average 2 percent cover. Creeping snowberry and thimbleberry are the most abundant and productive shrub species in this phase, and a diversity of other species are typically present. Other frequently occurring shrubs include Saskatoon serviceberry, Scoulers willow, bitter cherry, whitethorn ceanothus, Sierra currant, gooseberry currant, greenleaf manzanita, roundleaf snowberry, and Utah serviceberry. Antelope bitterbrush and mountain big sagebrush become more important with longer time intervals between fire. Forb richness is high, with over 40 species recorded in this community phase. More abundant forb species that frequently occur include rose thistle, woolly mules-ears, silvery lupine, and arrowleaf balsamroot. Squirreltail, western needlegrass (Achnatherum occidentale), and carex species (Carex spp.) are commonly occurring grasses or grasslike species.
Figure 8. Annual production by plant type (representative values) or group (midpoint values)
Table 5. Annual production by plant type
| Plant type | Low (kg/hectare) |
Representative value (kg/hectare) |
High (kg/hectare) |
|---|---|---|---|
| Shrub/Vine | 56 | 84 | 112 |
| Tree | 56 | 84 | 112 |
| Forb | 22 | 39 | 50 |
| Grass/Grasslike | 1 | 2 | 3 |
| Total | 135 | 209 | 277 |
Table 6. Soil surface cover
| Tree basal cover | 2-3% |
|---|---|
| Shrub/vine/liana basal cover | 0-1% |
| Grass/grasslike basal cover | 0% |
| Forb basal cover | 0-1% |
| Non-vascular plants | 0% |
| Biological crusts | 0% |
| Litter | 33-81% |
| Surface fragments >0.25" and <=3" | 0.5-15.0% |
| Surface fragments >3" | 9-13% |
| Bedrock | 0% |
| Water | 0% |
| Bare ground | 0.5-25.0% |
Table 7. Woody ground cover
| Downed wood, fine-small (<0.40" diameter; 1-hour fuels) | 2-3% |
|---|---|
| Downed wood, fine-medium (0.40-0.99" diameter; 10-hour fuels) | 2-3% |
| Downed wood, fine-large (1.00-2.99" diameter; 100-hour fuels) | 1-2% |
| Downed wood, coarse-small (3.00-8.99" diameter; 1,000-hour fuels) | 1-2% |
| Downed wood, coarse-large (>9.00" diameter; 10,000-hour fuels) | 1-2% |
| Tree snags** (hard***) | – |
| Tree snags** (soft***) | – |
| Tree snag count** (hard***) | |
| Tree snag count** (hard***) |
* Decomposition Classes: N - no or little integration with the soil surface; I - partial to nearly full integration with the soil surface.
** >10.16cm diameter at 1.3716m above ground and >1.8288m height--if less diameter OR height use applicable down wood type; for pinyon and juniper, use 0.3048m above ground.
*** Hard - tree is dead with most or all of bark intact; Soft - most of bark has sloughed off.
Table 8. Canopy structure (% cover)
| Height Above Ground (m) | Tree | Shrub/Vine | Grass/ Grasslike |
Forb |
|---|---|---|---|---|
| <0.15 | 0-1% | 5-15% | 0-5% | 1-6% |
| >0.15 <= 0.3 | 1-2% | 5-15% | 0-5% | 1-6% |
| >0.3 <= 0.6 | 3-6% | 5-20% | 0-3% | 3-14% |
| >0.6 <= 1.4 | 3-6% | 5-40% | – | 0-1% |
| >1.4 <= 4 | 3-6% | 0-6% | – | – |
| >4 <= 12 | 3-5% | – | – | – |
| >12 <= 24 | 6-25% | – | – | – |
| >24 <= 37 | 10-29% | – | – | – |
| >37 | – | – | – | – |
Community 1.4
Young forest infilling

Figure 9. Community Phase 1.4
This community phase is defined by a dense canopy and high basal area of mixed conifers, and dominance by white fir. Canopy cover ranges from 60 to 80 percent. Trees are overcrowded and often stressed due to competition for water and nutrients. This stress increases susceptibility to infection and death from pests and drought. Fire hazard is high in this community phase due to the deep accumulation of litter and coarse woody debris, dense standing dead and downed trees, and dense multi-layered structure of the forest.
Forest overstory. White fir dominates the forest canopy, which ranges from 60 to 80 percent cover. Jeffrey pine, red fir (at higher elevations), and incense cedar are secondary canopy species. Patches of quaking aspen may be present, although these will be shaded out if infilling continues. The overstory structure is multi-story, with dense stocking. Dominant trees are 60 to 120 feet tall, with diameters ranging from 10 to 30 inches.
Forest understory. Understory diversity is lower in this community phase. Shrubs average 10 percent cover, ranging from 1 to 17 percent. Forbs average 13 percent cover, ranging from 7 to 21 percent, and grass cover averages 3 percent, ranging from 0 to 5 percent. Shrub species frequently present include creeping snowberry, thimbleberry, Scoulers willow, Utah serviceberry, and Sierra currant. Western brackenfern (Pteridium aquilinum) is the most abundant herbaceous species in this community phase.
Table 9. Soil surface cover
| Tree basal cover | 4-6% |
|---|---|
| Shrub/vine/liana basal cover | 0-1% |
| Grass/grasslike basal cover | 0% |
| Forb basal cover | 0-1% |
| Non-vascular plants | 0% |
| Biological crusts | 0% |
| Litter | 44-60% |
| Surface fragments >0.25" and <=3" | 0-5% |
| Surface fragments >3" | 0.5-8.0% |
| Bedrock | 0% |
| Water | 0% |
| Bare ground | 0.5-10.0% |
Table 10. Woody ground cover
| Downed wood, fine-small (<0.40" diameter; 1-hour fuels) | 2-4% |
|---|---|
| Downed wood, fine-medium (0.40-0.99" diameter; 10-hour fuels) | 3-5% |
| Downed wood, fine-large (1.00-2.99" diameter; 100-hour fuels) | 5-10% |
| Downed wood, coarse-small (3.00-8.99" diameter; 1,000-hour fuels) | 5-15% |
| Downed wood, coarse-large (>9.00" diameter; 10,000-hour fuels) | 5-20% |
| Tree snags** (hard***) | – |
| Tree snags** (soft***) | – |
| Tree snag count** (hard***) | |
| Tree snag count** (hard***) |
* Decomposition Classes: N - no or little integration with the soil surface; I - partial to nearly full integration with the soil surface.
** >10.16cm diameter at 1.3716m above ground and >1.8288m height--if less diameter OR height use applicable down wood type; for pinyon and juniper, use 0.3048m above ground.
*** Hard - tree is dead with most or all of bark intact; Soft - most of bark has sloughed off.
Community 1.5
Old-growth forest infilling
The old-growth, closed white fir-mixed conifer forest develops with the continued exclusion of fire, causing tree density to increase to unhealthy levels. Competition for water and sunlight continue, and tree health and vigor declines. Canopy cover ranges from 60 to over 100 percent. Understory cover, production and diversity are low in this phase. The abundance of ladder and ground fuels makes this phase high-risk for high severity fire. Increased tree density also makes this phase more susceptible to insect outbreaks, which can increase mortality after fire or during drought.
Pathway 1.1a
Community 1.1 to 1.2
In the event of a severe canopy fire or a clear-cut the old-growth forest would transition to stand initiation (community phase 1.2).
Pathway 1.1b
Community 1.1 to 1.5
Occurs with long term fire suppression that leads to forest infilling (community phase 1.5).
Pathway 1.2a
Community 1.2 to 1.3
This pathway occurs with time, and once the young forest is established, with a natural fire regime with frequent low to medium severity fires occurring every 3 to 9 years. Manual thinning with prescribed burns can imitate the natural cycle and lead to the same young, open mixed-conifer forest (community phase 1.3).
Pathway 1.2b
Community 1.2 to 1.4
This pathway occurs when fire is excluded from the system, and leads to forest infilling with white fir, incense cedar and red fir increasing in the understory, and Jeffrey pine and sugar pine decreasing in importance (community phase 1.4).
Pathway 1.3a
Community 1.3 to 1.1
This pathway occurs with time with frequent low to moderate intensity burning. Manual thinning or prescribed burning can be implemented to replace the natural disturbances that keep this forest open. This pathway leads to community phase 1.1.
Pathway 1.3b
Community 1.3 to 1.2
In the event of a canopy fire this community phase would return to forest stand initiation (community phase 1.2).
Pathway 1.3c
Community 1.3 to 1.4
If fire does not occur, the density of the forest increases slowly over time, favoring white fir, incense cedar, and red fir over Jeffrey pine and sugar pine (community phase 1.4).
Pathway 1.4b
Community 1.4 to 1.2
The density of ground and ladder fuels creates conditions for a high intensity canopy fire. A severe fire would initiate stand regeneration (community phase 1.2). This can shift the community back to its natural state, but further treatments may be needed to eventually achieve the relatively open forest dominated by Jeffrey pine (community phase 1.3).
Pathway 1.4c
Community 1.4 to 1.3
A naturally occurring moderate or surface fire in this forest is unlikely due to the high fuel load. Considerable management efforts would be needed to create the open forest conditions that should exist in this forest under a natural fire regime. Manual treatment or prescribed burns could thin out fires and small incense cedar, as well as the fuels in the understory. This would shift this forest back to its natural state of a young, relatively open mixed conifer forest (community phase 1.3).
Pathway 1.4a
Community 1.4 to 1.5
If fire continues to be excluded from this system, the old-growth closed white fir-mixed conifer forest community develops (community phase 1.5).
Pathway 1.5b
Community 1.5 to 1.1
A naturally occurring moderate or surface fire in this forest is unlikely due to the high fuel load. Considerable management effort would be needed to create the relatively open forest conditions that should exist in this forest with a natural fire regime. Manual treatment with prescribed burns could thin out firs and incense cedar, as well as the fuels in the understory. This would shift this forest back to its natural state of a relatively open mixed conifer forest dominated by Jeffrey pine (community phase 1.1).
Pathway 1.5a
Community 1.5 to 1.2
At this point a severe fire is likely and would initiate stand regeneration (community phase 1.2).
Additional community tables
Table 11. Community 1.3 plant community composition
| Group | Common name | Symbol | Scientific name | Annual production (kg/hectare) | Foliar cover (%) | |
|---|---|---|---|---|---|---|
|
Tree
|
||||||
| 1 | Trees | 56–112 | ||||
| white fir | ABCO | Abies concolor | 50–106 | 8–30 | ||
| incense cedar | CADE27 | Calocedrus decurrens | 1–11 | 2–11 | ||
| Jeffrey pine | PIJE | Pinus jeffreyi | 3–8 | 6–31 | ||
| quaking aspen | POTR5 | Populus tremuloides | 0–6 | 0–20 | ||
| sugar pine | PILA | Pinus lambertiana | 0–6 | 0–3 | ||
|
Shrub/Vine
|
||||||
| 2 | Shrubs | 56–112 | ||||
| thimbleberry | RUPA | Rubus parviflorus | 0–56 | 0–20 | ||
| creeping snowberry | SYMO | Symphoricarpos mollis | 28–56 | 5–15 | ||
| whitethorn ceanothus | CECO | Ceanothus cordulatus | 0–22 | 0–7 | ||
| Saskatoon serviceberry | AMAL2 | Amelanchier alnifolia | 9–22 | 0–3 | ||
| antelope bitterbrush | PUTR2 | Purshia tridentata | 10–22 | 0–2 | ||
| bitter cherry | PREM | Prunus emarginata | 0–17 | 0–1 | ||
| mountain big sagebrush | ARTRV | Artemisia tridentata ssp. vaseyana | 0–13 | 0–3 | ||
| Utah serviceberry | AMUT | Amelanchier utahensis | 0–11 | 0–7 | ||
| greenleaf manzanita | ARPA6 | Arctostaphylos patula | 0–2 | 0–1 | ||
| gooseberry currant | RIMO2 | Ribes montigenum | 0–2 | 0–1 | ||
| Sierra currant | RINE | Ribes nevadense | 0–2 | 0–1 | ||
| roundleaf snowberry | SYRO | Symphoricarpos rotundifolius | 0–2 | 0–1 | ||
| Scouler's willow | SASC | Salix scouleriana | 0–2 | 0–1 | ||
|
Forb
|
||||||
| 3 | Forbs | 22–50 | ||||
| silvery lupine | LUAR3 | Lupinus argenteus | 0–28 | 0–5 | ||
| rose thistle | CIAN | Cirsium andersonii | 0–22 | 0–2 | ||
| woolly mule-ears | WYMO | Wyethia mollis | 0–11 | 0–5 | ||
| arrowleaf balsamroot | BASA3 | Balsamorhiza sagittata | 0–6 | 0–1 | ||
| wavyleaf Indian paintbrush | CAAP4 | Castilleja applegatei | 0–1 | 0–1 | ||
| dusky onion | ALCA2 | Allium campanulatum | 0–1 | 0–1 | ||
| spreading dogbane | APAN2 | Apocynum androsaemifolium | 0–1 | 0–1 | ||
| Holboell's rockcress | ARHO2 | Arabis holboellii | 0–1 | 0–1 | ||
| cryptantha | CRYPT | Cryptantha | 0–1 | 0–1 | ||
| sanddune wallflower | ERCAP | Erysimum capitatum var. perenne | 0–1 | 0–1 | ||
| milk kelloggia | KEGA | Kelloggia galioides | 0–1 | 0–1 | ||
| pea | LATHY | Lathyrus | 0–1 | 0–1 | ||
| Brown's peony | PABR | Paeonia brownii | 0–1 | 0–1 | ||
| goosefoot violet | VIPU4 | Viola purpurea | 0–1 | 0–1 | ||
|
Grass/Grasslike
|
||||||
| 4 | Grasses and Grasslike | 1–3 | ||||
| sedge | CAREX | Carex | 1–7 | 1–3 | ||
| western needlegrass | ACOC3 | Achnatherum occidentale | 0–4 | 0–2 | ||
| squirreltail | ELEL5 | Elymus elymoides | 0–1 | 0–1 | ||
Table 12. Community 1.3 forest overstory composition
| Common name | Symbol | Scientific name | Nativity | Height (m) | Canopy cover (%) | Diameter (cm) | Basal area (square m/hectare) |
|---|---|---|---|---|---|---|---|
|
Tree
|
|||||||
| Jeffrey pine | PIJE | Pinus jeffreyi | Native | – | 5–30 | 40.6–68.6 | – |
| white fir | ABCO | Abies concolor | Native | – | 7–28 | 48.3–68.6 | – |
| quaking aspen | POTR5 | Populus tremuloides | Native | – | 0–20 | – | – |
| incense cedar | CADE27 | Calocedrus decurrens | Native | – | 2–10 | – | – |
| sugar pine | PILA | Pinus lambertiana | Native | – | 0–3 | – | – |
Table 13. Community 1.3 forest understory composition
| Common name | Symbol | Scientific name | Nativity | Height (m) | Canopy cover (%) | |
|---|---|---|---|---|---|---|
|
Grass/grass-like (Graminoids)
|
||||||
| sedge | CAREX | Carex | Native | – | 1–3 | |
| western needlegrass | ACOC3 | Achnatherum occidentale | Native | – | 0–2 | |
| squirreltail | ELEL5 | Elymus elymoides | Native | – | 0–1 | |
|
Forb/Herb
|
||||||
| silvery lupine | LUAR3 | Lupinus argenteus | Native | – | 0–5 | |
| woolly mule-ears | WYMO | Wyethia mollis | Native | – | 0–5 | |
| rose thistle | CIAN | Cirsium andersonii | Native | – | 0–2 | |
| spreading dogbane | APAN2 | Apocynum androsaemifolium | Native | – | 0–1 | |
| sanddune wallflower | ERCAP | Erysimum capitatum var. perenne | Native | – | 0–1 | |
| Brown's peony | PABR | Paeonia brownii | Native | – | 0–1 | |
| pea | LATHY | Lathyrus | Native | – | 0–1 | |
| goosefoot violet | VIPU4 | Viola purpurea | Native | – | 0–1 | |
| arrowleaf balsamroot | BASA3 | Balsamorhiza sagittata | Native | – | 0–1 | |
| Holboell's rockcress | ARHO2 | Arabis holboellii | Native | – | 0–1 | |
| milk kelloggia | KEGA | Kelloggia galioides | Native | – | 0–1 | |
| wavyleaf Indian paintbrush | CAAP4 | Castilleja applegatei | Native | – | 0–1 | |
| cryptantha | CRYPT | Cryptantha | Native | – | 0–1 | |
| dusky onion | ALCA2 | Allium campanulatum | Native | – | 0–1 | |
|
Shrub/Subshrub
|
||||||
| thimbleberry | RUPA | Rubus parviflorus | Native | – | 1–20 | |
| creeping snowberry | SYMO | Symphoricarpos mollis | Native | – | 5–15 | |
| Utah serviceberry | AMUT | Amelanchier utahensis | Native | – | 0–7 | |
| whitethorn ceanothus | CECO | Ceanothus cordulatus | Native | – | 0–7 | |
| mountain big sagebrush | ARTRV | Artemisia tridentata ssp. vaseyana | Native | – | 0–3 | |
| Saskatoon serviceberry | AMAL2 | Amelanchier alnifolia | Native | – | 0–3 | |
| antelope bitterbrush | PUTR2 | Purshia tridentata | Native | – | 0.5–2 | |
| bitter cherry | PREM | Prunus emarginata | Native | – | 0–1 | |
| Scouler's willow | SASC | Salix scouleriana | Native | – | 0–1 | |
| gooseberry currant | RIMO2 | Ribes montigenum | Native | – | 0–1 | |
| greenleaf manzanita | ARPA6 | Arctostaphylos patula | Native | – | 0–1 | |
| roundleaf snowberry | SYRO | Symphoricarpos rotundifolius | Native | – | 0–1 | |
| Sierra currant | RINE | Ribes nevadense | Native | – | 0–1 | |
|
Tree
|
||||||
| white fir | ABCO | Abies concolor | Native | – | 0.5–2 | |
| sugar pine | PILA | Pinus lambertiana | Native | – | 0–2 | |
| quaking aspen | POTR5 | Populus tremuloides | Native | – | 0–2 | |
| incense cedar | CADE27 | Calocedrus decurrens | Native | – | 0.5–1 | |
| Jeffrey pine | PIJE | Pinus jeffreyi | Native | – | 0.5–1 | |
Table 14. Community 1.4 forest overstory composition
| Common name | Symbol | Scientific name | Nativity | Height (m) | Canopy cover (%) | Diameter (cm) | Basal area (square m/hectare) |
|---|---|---|---|---|---|---|---|
|
Tree
|
|||||||
| white fir | ABCO | Abies concolor | Native | – | 17–55 | 25.4–76.2 | – |
| quaking aspen | POTR5 | Populus tremuloides | Native | – | 0–20 | – | – |
| California red fir | ABMA | Abies magnifica | Native | – | 0–10 | – | – |
| incense cedar | CADE27 | Calocedrus decurrens | Native | – | 0–10 | – | – |
| Jeffrey pine | PIJE | Pinus jeffreyi | Native | – | 3–8 | – | – |
Table 15. Community 1.4 forest understory composition
| Common name | Symbol | Scientific name | Nativity | Height (m) | Canopy cover (%) | |
|---|---|---|---|---|---|---|
|
Grass/grass-like (Graminoids)
|
||||||
| needlegrass | ACHNA | Achnatherum | Native | – | 0–3 | |
| big quakinggrass | BRMA | Briza maxima | Native | – | 0–1 | |
|
Forb/Herb
|
||||||
| longspur lupine | LUAR6 | Lupinus arbustus | Native | – | 0–2 | |
| waxy checkerbloom | SIGL2 | Sidalcea glaucescens | Native | – | 0–1 | |
| white hawkweed | HIAL2 | Hieracium albiflorum | Native | – | 0–1 | |
| Gray's licorice-root | LIGR | Ligusticum grayi | Native | – | 0–0.5 | |
| Virginia strawberry | FRVI | Fragaria virginiana | Native | – | 0–0.5 | |
| milk kelloggia | KEGA | Kelloggia galioides | Native | – | 0–0.5 | |
| common yarrow | ACMI2 | Achillea millefolium | Native | – | 0–0.5 | |
|
Fern/fern ally
|
||||||
| western brackenfern | PTAQ | Pteridium aquilinum | Native | – | 0–4 | |
|
Shrub/Subshrub
|
||||||
| creeping snowberry | SYMO | Symphoricarpos mollis | Native | – | 0–6 | |
| thimbleberry | RUPA | Rubus parviflorus | Native | – | 0.5–2 | |
| Utah serviceberry | AMUT | Amelanchier utahensis | Native | – | 0–2 | |
| Scouler's willow | SASC | Salix scouleriana | Native | – | 0–1 | |
| Sierra currant | RINE | Ribes nevadense | Native | – | 0–1 | |
|
Tree
|
||||||
| white fir | ABCO | Abies concolor | Native | – | 3–5 | |
| California red fir | ABMA | Abies magnifica | Native | – | 0–3 | |
| quaking aspen | POTR5 | Populus tremuloides | Native | – | 0–2 | |
Interpretations
Animal community
This forest provides food and shelter for a variety of animals including squirrels, bears, birds and deer. Tree seeds are eaten by birds, and the roots and young stems are eaten by small mammals. The standing dead and downed trees provide habitats for nesting birds and shelter for cavity dwellers (Gucker 2007).
Thimbleberry is an important food source for deer and elk, black bear, small mammals and birds (Gucker 2012).
Hydrological functions
The hydrology of this site is characterized by heavy snowmelt in the spring, with very little precipitation in the summer months.
Recreational uses
This ecological site is can be a scenic forest. If slopes are appropriate, it provides suitable camping and picnicking areas. Trails for walking, biking and cross-country skiing are found along the shore of Lake Tahoe, and upland throughout this site.
Wood products
Jeffrey pine is used for high-grade lumber, and is used for molding, cabinets, doors, and windows (Gucker 2007).
White fir is used extensively for solid construction framing and plywood, pulpwood, poles and pilings. It requires large amounts of preservatives because the heartwood decays rapidly. It is poorly suited for firewood because of its low specific gravity and heat production (Zouhar 2001).
Sugar pine lumber is valued because it is easy to mill and can be used for molding, window and doorframes, doors and other specialty items (Habeck 1992).
The wood from incense cedar is valued because is resistant to rot. It is used as mud sills, window sashes, sheathing under stucco or brick veneer construction, greenhouse benches, fencing, poles, and trellises. It is also widely used for exterior and interior siding. Much of the top quality incense cedar is used in the manufacture of pencils (Tollefson 2008).
Other products
Jeffrey pine and sugar pine cones are used for arts and crafts.
Jeffrey pine pitch was distilled for turpentine early in the century; however the terpenes were found to contain high amounts of the explosive chemical heptane (Gucker 2007).
Other information
Thimbleberry produces delicious fruits that were widely consumed by many indigenous peoples, and stems and leaves were used for medicinal purposes and in basketry (Gucker 2012).
Table 16. Representative site productivity
Supporting information
Inventory data references
Community Phase 1.1:
JhC74001 - Type location
tcb03012
tcc03010
Community Phase 1.4:
tcc03063
tcc03064
tkc03066
Type locality
| Location 1: Placer County, CA | |
|---|---|
| Township/Range/Section | T16N R17E S12 |
| UTM zone | N |
| UTM northing | 48123 |
| UTM easting | 55116 |
| General legal description | Approximately 0.5 miles west of highway 267 in Kings Beach, CA. |
Other references
Anderson, M. D. 2001. Salix scouleriana. Fire Effects Information System, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory.
Anderson, M. D. 2008. Carex rossii. Fire Effects Information System, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory.
Azuma, D. L., J. Donnegan, and D. Gedney. 2004. Southwest Oregon Biscuit Fire: an analysis of forest resources and fire severity. PNW-RP-560, USDA Forest Service, Pacific Northwest Research Station.
Barbour, M., E. Kelly, P. Maloney, D. Rizzo, E. Royce, and J. Fites-Kaufmann. 2002. Present and past old-growth forests of the Lake Tahoe Basin, Sierra Nevada, US. Journal of Vegetation Science 13:461-472.
Beardsley, D., C. Bolsinger, and R. Warbington. 1999. Old-growth forest in the Sierra Nevada: By type in 1945 and 1993 and ownership in 1999. PNW-RP-516, USDA Forest Service, Pacific Northwest Research Station.
Beaty, R. M. and A. H. Taylor. 2008. Fire history and the structure and dynamics of a mixed conifer forest landscape in the northern Sierra Nevada, Lake Tahoe Basin, California, USA. Forest Ecology and Management 255:707-719.
Binkley, D., T. Sisk, C. Chambers, J. Springer, and W. Block. 2007. The role of old-growth forests in frequent-fire landscapes. Ecology and Society 12:18-35.
Bradley, T. and P. Tueller. 2001. Effects of fire on bark beetle presence on Jeffrey pine in the Lake Tahoe Basin. Forest Ecology and Management 142:205-214.
Dhaemers. 2006. Vegetation recovery following spring prescribed fire in pinyon-juniper woodlands of central Nevada: Effects of elevation and tree cover. M.Sc. Unversity of Nevada, Reno.
Elliot-Fisk, D. L., R. Harris, R. A. Rowntree, T. C. Cahill, R. Kattelmann, P. Rucks, O. K. Davis, R. Lacey, D. A. Sharkey, L. Duan, D. Leisz, S. L. Stephens, C. R. Goldman, S. Lindstrom, D. S. Ziegler, G. E. Gruell, and D. Machida. 1996. Lake Tahoe Case Study. Pages 217-276 Sierra Nevada Ecosystem Project. University of California, Centers for Water and Wildland Resources, Davis, CA.
Fettig, C. J., S. R. McKelvey, D. R. Cluck, S. L. Smith, and W. J. Otrosina. 2010. Effects of prescribed fire and season of burn on direct and indirect levels of tree mortality in Ponderosa and Jeffrey Pine forests in California, USA. Forest Ecology and Management 260:207-218.
Fryer, J. L. 1997. Amelanchier alnifolia. Fire Effects Information System, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory.
Gucker, C. 2012. Rubus parviflorus. Fire Effects Information System, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory.
Habeck, R. J. 1992. Pinus lambertiana. Fire Effects Information System, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory.
Hauser, A. S. 2007. Arctostaphylos patula. Fire Effects Information System, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory.
Huisinga, K. D., D. C. Laughlin, P. Z. Fule, J. D. Springer, and C. M. McGlone. 2005. Effects of an intense prescribed fire on understory vegetation in a mixed conifer forest. Journal of the Torrey Botanical Society 132:590-601.
Jones, M. E., T. D. Paine, M. E. Fenn, and M. A. Poth. 2004. Influence of ozone and nitrogen deposition on bark beetle activity under drought conditions. Forest Ecology and Management 200:67-76.
Laughlin, D. C., J. D. Bakker, and P. Z. Fule. 2005. Understorey plant community structure in lower montane and subalpine forests, Grand Canyon National Park, USA. Journal of Biogeography 32:2083-2102.
Lindh, B. C. and P. S. Muir. 2004. Understory vegetation in young Douglas-fir forests: does thinning help restore old-growth composition? Forest Ecology and Management 192:285-296.
Maloney, P. E., T. F. Smith, C. E. Jensen, J. Innes, D. M. Rizzo, and M. P. North. 2008. Initial tree mortality and insect and pathogen response to fire and thinning restoration treatments in an old-growth mixed-conifer forest of the Sierra Nevada, California. Canadian Journal of Forest Research 38:3011-3020.
Murphy, D. D. and C. M. Knopp. 2000. Lake Tahoe Basin Watershed Assessment. PSW-GTR-175, USDA Forest Service, Pacific Southwest Research Station.
Nagal, T. A. and A. H. Taylor. 2005. Fire and persistence of montane chaparral in mixed conifer forest landscapes in the northern Sierra Nevada, Lake Tahoe Basin, California, USA. Journal of the Torrey Botanical Society 132:442-457.
Parker, V. T. and M. P. Yoder-Williams. 1989. Reduction of survival and growth of young Pinus jeffreyi by an herbaceous perennial, Wyethia mollis. American Midland Naturalist 121:105-111.
Peterson, D. L., M. J. Arbaugh, V. A. Wakefield, and P. R. Miller. 1987. Evidence of growth reduction in Ozone-injured Jeffrey pine (Pinus jeffreyi Grev. and Balf.) in Sequoia and Kings Canyon National Parks. JAPCA 37:906-912.
Rau, B. M., J. C. Chambers, R. R. Blank, and D. W. Johnson. 2008. Prescribed fire, soil, and plants: burn effects and interactions in the central Great Basin. Rangeland Ecology and Management 61:169-181.
Reeves, S. L. 2006. Ceanothus cordulatus. Fire Effects Information System. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory.
Riegel, G. M., T. J. Svejcar, and M. D. Busse. 2002. Does the presence of Wyethia mollis affect growth of Pinus jeffreyi seedlings? Western North American Naturalist 62:141-150.
Schoennagel, T., D. M. Waller, M. G. Turner, and W. H. Romme. 2004. The effect of fire interval on post-fire understorey communities in Yellowstone National Park. Journal of Vegetation Science 15:797-806.
Simonin, K. A. 2001. Elymus elymoides. Fire Effects Information System, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory.
Smith, S. 1994. Ecological guide to eastside pine associations. R5-ECOL-TP-004, USDA Forest Service, Pacific Southwest Region.
Snyder, S. A. 1991. Symphoricarpos mollis. Fire Effects Information System, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory.
Stephens, S. L. and D. L. Fry. 2005. Spatial distribution of regeneration patches in an old-growth Pinus jeffrey-mixed conifer forest in northwestern Mexico. Journal of Vegetation Science 16:693-702.
Taylor, E. H. 2004. Identifying forest reference conditions on early cut-over lands, Lake Tahoe Basin, USA. Ecological Applications 14:1903-1920.
Tollefson, J. E. 2008. Calocedrus decurrens. Fire Effects Information System, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory.
Wayman, R. B. and M. North. 2007. Initial response of a mixed-conifer understory plant community to burning and thinning restoration treatments. Forest Ecology and Management 239:32-44.
Wright, H. A. 1985. Effects of fire on grasses and forbs in sagebrush-grass communities. Pages 12-21 in Rangeland Fire Effects; A symposium. USDI-Bureau of Land Management, Boise, ID.
Zhang, J., M. W. Ritchie, and W. W. Oliver. 2008. Vegetation responses to stand structure and prescribed fire in an interior ponderosa pine ecosystem. Canadian Journal of Forest Research 38:909-918.
Zouhar, K. 2001. Abies concolor. Fire Effects Information System, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory.
Contributors
Alice Miller
Marchel Munnecke
Rangeland health reference sheet
Interpreting Indicators of Rangeland Health is a qualitative assessment protocol used to determine ecosystem condition based on benchmark characteristics described in the Reference Sheet. A suite of 17 (or more) indicators are typically considered in an assessment. The ecological site(s) representative of an assessment location must be known prior to applying the protocol and must be verified based on soils and climate. Current plant community cannot be used to identify the ecological site.
| Author(s)/participant(s) | |
|---|---|
| Contact for lead author | |
| Date | |
| Approved by | |
| Approval date | |
| Composition (Indicators 10 and 12) based on | Annual Production |
Indicators
-
Number and extent of rills:
-
Presence of water flow patterns:
-
Number and height of erosional pedestals or terracettes:
-
Bare ground from Ecological Site Description or other studies (rock, litter, lichen, moss, plant canopy are not bare ground):
-
Number of gullies and erosion associated with gullies:
-
Extent of wind scoured, blowouts and/or depositional areas:
-
Amount of litter movement (describe size and distance expected to travel):
-
Soil surface (top few mm) resistance to erosion (stability values are averages - most sites will show a range of values):
-
Soil surface structure and SOM content (include type of structure and A-horizon color and thickness):
-
Effect of community phase composition (relative proportion of different functional groups) and spatial distribution on infiltration and runoff:
-
Presence and thickness of compaction layer (usually none; describe soil profile features which may be mistaken for compaction on this site):
-
Functional/Structural Groups (list in order of descending dominance by above-ground annual-production or live foliar cover using symbols: >>, >, = to indicate much greater than, greater than, and equal to):
Dominant:
Sub-dominant:
Other:
Additional:
-
Amount of plant mortality and decadence (include which functional groups are expected to show mortality or decadence):
-
Average percent litter cover (%) and depth ( in):
-
Expected annual annual-production (this is TOTAL above-ground annual-production, not just forage annual-production):
-
Potential invasive (including noxious) species (native and non-native). List species which BOTH characterize degraded states and have the potential to become a dominant or co-dominant species on the ecological site if their future establishment and growth is not actively controlled by management interventions. Species that become dominant for only one to several years (e.g., short-term response to drought or wildfire) are not invasive plants. Note that unlike other indicators, we are describing what is NOT expected in the reference state for the ecological site:
-
Perennial plant reproductive capability:
Print Options
Sections
Font
Other
The Ecosystem Dynamics Interpretive Tool is an information system framework developed by the USDA-ARS Jornada Experimental Range, USDA Natural Resources Conservation Service, and New Mexico State University.
Click on box and path labels to scroll to the respective text.