Natural Resources
Conservation Service
Ecological site R026XY021NV
SODIC FLAT
Last updated: 4/10/2024
Accessed: 03/10/2026
General information
Provisional. A provisional ecological site description has undergone quality control and quality assurance review. It contains a working state and transition model and enough information to identify the ecological site.
MLRA notes
Major Land Resource Area (MLRA): 026X–Carson Basin and Mountains
The area lies within western Nevada and eastern California, with about 69 percent being within Nevada, and 31 percent being within California. Almost all this area is in the Great Basin Section of the Basin and Range Province of the Intermontane Plateaus. Isolated north-south trending mountain ranges are separated by aggraded desert plains. The mountains are uplifted fault blocks with steep side slopes. Most of the valleys are drained by three major rivers flowing east across this MLRA. A narrow strip along the western border of the area is in the Sierra Nevada Section of the Cascade-Sierra Mountains Province of the Pacific Mountain System. The Sierra Nevada Mountains are primarily a large fault block that has been uplifted with a dominant tilt to the west. This structure leaves an impressive wall of mountains directly west of this area. This helps create a rain shadow affect to MLRA 26. Parts of this eastern face, but mostly just the foothills, mark the western boundary of this area. Elevations range from about 3,806 feet (1,160 meters) on the west shore of Pyramid Lake to 11,653 feet (3,552 meters) on the summit of Mount Patterson in the Sweetwater Mountains.
Valley areas are dominantly composed of Quaternary alluvial deposits with Quaternary playa or alluvial flat deposits often occupying the lowest valley bottoms in the internally drained valleys, and river deposited alluvium being dominant in externally drained valleys. Hills and mountains are dominantly Tertiary andesitic flows, breccias, ash flow tuffs, rhyolite tuffs or granodioritic rocks. Quaternary basalt flows are present in lesser amounts, and Jurassic and Triassic limestone and shale, and Precambrian limestone and dolomite are also present in very limited amounts. Also of limited extent are glacial till deposits along the east flank of the Sierra Nevada Mountains, the result of alpine glaciation.
The average annual precipitation in this area is 5 to 36 inches (125 to 915 millimeters), increasing with elevation. Most of the rainfall occurs as high-intensity, convective storms in spring and autumn. Precipitation is mostly snow in winter. Summers are dry. The average annual temperature is 37 to 54 degrees F (3 to 12 degrees C). The freeze-free period averages 115 days and ranges from 40 to 195 days, decreasing in length with elevation.
The dominant soil orders in this MLRA are Aridisols and Mollisols. The soils in the area dominantly have a mesic soil temperature regime, an aridic or xeric soil moisture regime, and mixed or smectitic mineralogy. They generally are well drained, are clayey or loamy and commonly skeletal, and are very shallow to moderately deep.
This area supports shrub-grass vegetation characterized by big sagebrush. Low sagebrush and Lahontan sagebrush occur on some soils. Antelope bitterbrush, squirreltail, desert needlegrass, Thurber needlegrass, and Indian ricegrass are important associated plants. Green ephedra, Sandberg bluegrass, Anderson peachbrush, and several forb species also are common. Juniper-pinyon woodland is typical on mountain slopes. Jeffrey pine, lodgepole pine, white fir, and manzanita grow on the highest mountain slopes. Shadscale is the typical plant in the drier parts of the area. Sedges, rushes, and moisture-loving grasses grow on the wettest parts of the wet flood plains and terraces. Basin wildrye, alkali sacaton, saltgrass, buffaloberry, black greasewood, and rubber rabbitbrush grow on the drier sites that have a high concentration of salts.
Some of the major wildlife species in this area are mule deer, coyote, beaver, muskrat, jackrabbit, cottontail, raptors, pheasant, chukar, blue grouse, mountain quail, and mourning dove. The species of fish in the area include trout and catfish. The Lahontan cutthroat trout in the Truckee River is a threatened and endangered species.
LRU notes
The Semiarid Fans and Basins LRU includes basins, alluvial fans and adjacent hill slopes immediately east of the Sierra Nevada mountain range and are affected by its climate or have its granitic substrate. Elevations range from 1355 to 1920 meters and slopes range from 0 to 30 percent, with a median value of 6 percent. Frost free days range from 121 to 170.
Ecological site concept
The Sodic Flat site occurs on lake terraces and flood plains. Slopes range from 0 to 2 percent. Elevations are 4200 to 5500 feet. The soils associated with this site are very deep, moderately well drained soils that formed in alluvium over lacustrine deposits derived from mixed rock sources. They have a surface layer that tends to be strongly sodium affected. This surface layer will crust and bake upon drying. These factors will inhibit water infiltration and seedling emergence. Endosaturation is present with an apparent seasonal high water table that fluctuates between 125 and 175 cm between February and May. Greasewood (Sarcobatus vermiculatus) can take advantage of the deep water table while shallow rooted plants cannot. Flooding is minimal, although some areas are subject to ponding for brief periods due mainly to run-in from adjacent areas.
Associated sites
| R026XY001NV |
MOIST FLOODPLAIN |
|---|---|
| R026XY002NV |
WET SODIC BOTTOM |
| R026XY004NV |
SALINE BOTTOM |
| R026XY013NV |
SODIC FLOODPLAIN |
Similar sites
| R026XY002NV |
WET SODIC BOTTOM DISP dominant plant; more productive site |
|---|---|
| R026XY004NV |
SALINE BOTTOM LECI2 dominant plant on site; more productive site |
| R026XY013NV |
SODIC FLOODPLAIN SPAI dominant grass; more productive site |
Table 1. Dominant plant species
| Tree |
Not specified |
|---|---|
| Shrub |
(1) Sarcobatus vermiculatus |
| Herbaceous |
(1) Leymus cinereus |
Physiographic features
The Sodic Flat site occurs on lake terraces and flood plains. Slopes range from 0 to 2 percent. Elevations are 4200 to 5500 feet.
Table 2. Representative physiographic features
| Landforms |
(1)
Lake terrace
(2) Flood plain |
|---|---|
| Flooding duration | Very brief (4 to 48 hours) to brief (2 to 7 days) |
| Flooding frequency | None to occasional |
| Ponding frequency | None |
| Elevation | 1,280 – 1,676 m |
| Slope | 0 – 2% |
| Water table depth | 91 – 183 cm |
| Aspect | Aspect is not a significant factor |
Climatic features
The climate is semiarid with cool, moist winters and warm, dry summers. Average annual precipitation is 8 to 10 inches. Mean annual air temperature is 45 to 50 degrees F. The average growing season is about 80 to 110 days.
Average annual precipitation (in inches) by month at the Stead climate station is: Jan. 1.57, Feb. 1.92, Mar. 1.24, Apr. 0.56, May 0.55, Jun. 0.54, Jul. 0.34, Aug. 0.26, Sep. 0.46, Oct. 0.72, Nov. 0.88, Dec. 1.84. Average annual precipitation is 10.89 inches.
Nevada’s climate is predominantly arid, with large daily ranges of temperature, infrequent severe storms, heavy snowfall in the higher mountains, and great location variations with elevation. Three basic geographical factors largely influence Nevada’s climate: continentality, latitude, and elevation. Continentality is the most important factor. The strong continental effect is expressed in the form of both dryness and large temperature variations. Nevada lies on the eastern, lee side of the Sierra Nevada Range, a massive mountain barrier that markedly influences the climate of the State. The prevailing winds are from the west, and as the warm moist air from the Pacific Ocean ascend the western slopes of the Sierra Range, the air cools, condensation occurs and most of the moisture falls as precipitation. As the air descends the eastern slope, it is warmed by compression, and very little precipitation occurs. The effects of this mountain barrier are felt not only in the West but throughout the state, with the result that the lowlands of Nevada are largely desert or steppes. The temperature regime is also affected by the blocking of the inland-moving maritime air. Nevada sheltered from maritime winds, has a continental climate with well-developed seasons and the terrain responds quickly to changes in solar heating.
Nevada lies within the mid-latitude belt of prevailing westerly winds which occur most of the year. These winds bring frequent changes in weather during the late fall, winter and spring months, when most of the precipitation occurs. To the south of the mid-latitude westerlies, lies a zone of high pressure in subtropical latitudes, with a center over the Pacific Ocean. In the summer, this high-pressure belt shifts northward over the latitudes of Nevada, blocking storms from the ocean. The resulting weather is mostly clear and dry during the summer and early fall, with scattered thundershowers. The eastern portion of the state receives significant summer thunderstorms generated from monsoonal moisture pushed up from the Gulf of California, known as the North American monsoon. The monsoon system peaks in August and by October the monsoon high over the Western U.S. begins to weaken and the precipitation retreats southward towards the tropics (NOAA 2004).
Table 3. Representative climatic features
| Frost-free period (characteristic range) | |
|---|---|
| Freeze-free period (characteristic range) | |
| Precipitation total (characteristic range) | 203-254 mm |
| Frost-free period (average) | 95 days |
| Freeze-free period (average) | |
| Precipitation total (average) | 229 mm |
Figure 1. Monthly precipitation range
Figure 2. Monthly average minimum and maximum temperature
Figure 3. Annual precipitation pattern
Figure 4. Annual average temperature pattern
Influencing water features
Flooding is minimal, although some areas are subject to ponding for brief periods due mainly to run-in from adjacent areas.
Soil features
The soils associated with this site are very deep, moderately well drained soils that formed in alluvium over lacustrine deposits derived from mixed rock sources. They have a surface layer that tends to be strongly sodium affected. This surface layer will crust and bake upon drying. These factors will inhibit water infiltration and seedling emergence. Endosaturation is present with an apparent seasonal high water table that fluctuates between 125 and 175 cm between February and May. Greasewood can take advantage of the deep water table while shallow rooted plants cannot. Flooding is minimal, although some areas are subject to ponding for brief periods due mainly to run-in from adjacent areas. Runoff is low to very high. The soil moisture regime is aridic bordering on xeric and the soil temperature regime is mesic. Sheet and rill erosion is slight. Soil series associated with this site include: Doten, Mellor, Updike, and Vamp.
Table 4. Representative soil features
| Surface texture |
(1) Loam |
|---|---|
| Family particle size |
(1) Clayey |
| Drainage class | Moderately well drained |
| Permeability class | Slow |
| Soil depth | 183 – 213 cm |
| Surface fragment cover <=3" | 0% |
| Surface fragment cover >3" | 0% |
| Available water capacity (0-101.6cm) |
10.92 – 16.51 cm |
| Calcium carbonate equivalent (0-101.6cm) |
0% |
| Electrical conductivity (0-101.6cm) |
0 – 32 mmhos/cm |
| Sodium adsorption ratio (0-101.6cm) |
13 – 45 |
| Soil reaction (1:1 water) (0-101.6cm) |
8.2 – 9.6 |
| Subsurface fragment volume <=3" (Depth not specified) |
0% |
| Subsurface fragment volume >3" (Depth not specified) |
0% |
Ecological dynamics
An ecological site is the product of all the environmental factors responsible for its development and it has a set of key characteristics that influence a site’s resilience to disturbance and resistance to invasive species. Key characteristics include 1) climate (precipitation, temperature), 2) topography (aspect, slope, elevation, and landform), 3) hydrology (infiltration, runoff), 4) soils (depth, texture, structure, organic matter, salt content), 5) plant communities (functional groups, productivity), and 6) natural disturbance regime (fire, herbivory, etc.) (Caudle et al 2013). Biotic factors that that influence resilience include site productivity, species composition and structure, and population regulation and regeneration (Chambers et al. 2013).
Periodic drought regularly influences sagebrush ecosystems, and drought duration and severity has increased throughout the 20th century in much of the Intermountain West. Major shifts away from historic precipitation patterns have the greatest potential to alter ecosystem function and productivity (Snyder et al. 2019). Species composition and productivity can be altered by the timing of precipitation and water availability within the soil profile (Bates et al. 2006). The invasibility of plant communities is often linked to resource availability. Disturbance can decrease resource uptake due to damage or mortality of the native species and depressed competition or can increase resource pools by the decomposition of dead plant material following disturbance.
Black greasewood is classified as a phreatophyte, meaning it relies on access to groundwater rather than precipitation for survival (Meinzer 1927, Eddleman 2002, Naumberg et al. 2005). Its distribution is well correlated with the distribution of shallow groundwater (Meinzer 1927, Mozingo 1987). Black greasewood stands develop best where moisture is readily available, either from surface or subsurface runoff (Brown 1971). It is commonly found on floodplains that are either subjected to periodic flooding, have a high water table at least part of the year, or have a water table less than 34 feet deep (Harr and Price 1972, Blauer et al. 1976, Branson et al. 1976, Blaisdell and Holmgren 1984, Eddleman 2002). Romo (1984) found water tables ranging from 3.5 to 15 m under black greasewood dominated communities in Oregon. Black greasewood is usually a deep-rooted shrub, but has some shallow roots near the soil surface; the maximum rooting depth can be determined by the depth to a saturated zone (Harr and Price 1972).
Black greasewood is adapted to areas with shallow groundwater: it is tolerant of seasonal inundation but not continuous flooding, and its roots must have access to the water table for long-term survival. Ganskopp (1986) reported that water tables within 9.8 to 11.8 inches of the surface had no negative effect on black greasewood in Oregon. However, a study, conducted in California, found that black greasewood did not survive six months of continuous flooding (Groeneveld and Crowley 1988, Groeneveld 1990).
Black greasewood is capable of rooting to exceptional depths to access the water table in drought conditions. The taproots of black greasewood can penetrate from 20 to 57 feet below the surface (Meinzer 1927). However, other studies have found the maximum rooting depth to be closer to 12 feet (3.6 m), with effective use of water occurring at even shallower depths (Groeneveld 1990). Disturbances such as long-term drought and groundwater extraction that lower the water table beyond the rooting depths of these plants threaten communities of phreatophytic vegetation (Naumberg et al. 2005, Elmore et al. 2006). Recent remote sensing research in black greasewood and saltgrass communities shows a reduction in plant productivity over time with lowered water tables associated with groundwater pumping (Huntington et al. 2016). The exact groundwater level at which greasewood can no longer survive is not yet known (Devitt and Bird 2016). Lowering of the water table and subsequent loss of greasewood has been observed in other MLRAs. Death of phreatophytes in this system leaves the site open to invasion by non-native species (Devitt and Bird 2016, Provencher et al. 2020). Because of the high salt content of these soils, other more drought-tolerant native plants such as basin big sagebrush may be unlikely to colonize the site.
Basin wildrye is the dominant grass on this site. It is weakly rhizomatous and has been found to root to depths of up to 2 meters, and exhibits greater lateral root spread than many other grass species (Abbott et al. 1991, Reynolds and Fraley 1989). Basin wildrye is a large, cool-season perennial bunchgrass with an extensive deep coarse fibrous root system (Reynolds and Fraley 1989). Clumps may reach up to six feet in height (Ogle et al. 2012b). Basin wildrye does not tolerate long periods of inundation; it prefers cycles of wet winters and dry summers and is most commonly found in deep soils with high water holding capacities or seasonally high water tables (Ogle et al. 2012b, Perryman and Skinner 2007).
Seasonally high water tables have been found to be necessary for maintenance of site productivity and reestablishment of basin wildrye stands following disturbances such as fire, drought or excessive herbivory (Eckert et al. 1973). The sensitivity of basin wildrye seedling establishment to reduced soil water availability is increased as soil pH increases (Stuart et al. 1971). Lowering of the water table through extended drought, channel incision or groundwater pumping will decrease basin wildrye production and establishment, while sagebrush, rabbitbrush, and invasive weeds increase.
Other grasses that occur on this site are warm-season grasses, which means they use the C4 photosynthetic pathway. This adaptation makes these plants more efficient in their use of nitrogen and water (Taylor et al. 2009). These types of grasses can adjust their growth more quickly to drought or wet conditions when compared to C3 grasses (Witwicki et al. 2016).
Inland saltgrass is a warm-season rhizomatous grass that is often indicative of shallow groundwater. It is tolerant of high concentrations of salt (Skougard et al. 1979). Where present, the water table tends to be within 8 to 12 feet of the soil surface even in dry periods (Meinzer 1927). Saltgrass is also adapted to low water conditions, as it can distribute water for long distances through its connected rhizomes (Alpert 1990).
Alkali sacaton, a minor component of this site, is a native, long-lived, warm-season, densely tufted perennial bunchgrass ranging from 20 to 40 inches in height. It usually grows on saline soils but is not restricted to saline soils and can be found on nonsaline soils, rocky sites, open plains, valleys and bottom lands (Dayton 1937). Marcum and Kopec (1997) found inland saltgrass more tolerant of increased levels of salinity than alkali sacaton, therefore dewatering and/or long term drought that cause increased levels of salinity would create environmental conditions more favorable to inland saltgrass over alkali sacaton. Alkali sacaton is considered a facultative species in this region; it is tolerant of drought and inundation (Brakie 2007).
These communities often exhibit the formation of microbiotic crusts within the interspaces. These crusts influence the soils on these sites and their ability to reduce erosion and increase infiltration, they may also alter the soil structure and possibly increase soil fertility (Fletcher and Martin 1948, Williams 1993). Finer textured soils such as silts tend to support more microbiotic cover than coarse textured soils (Anderson et al. 1982). Disturbance such as hoof action from inappropriate grazing and cheatgrass (Bromus tectorum) invasion can reduce biotic crust integrity (Anderson et al. 1982, Ponzetti et al. 2007) and increase erosion. Annual non-native species such as clasping pepperweed (Lepidium perfoliatum), crossflower (aka purple mustard, Chorispora tenella), bur buttercup (Ceratocephela testiculata), and cheatgrass invade these sites where competition from perennial species is decreased. Native annual forbs like western tansymustard (Descurainia pinnata) can also become weedy on this site. Density of western tansymustard increases with supplemental water (Gutierrez and Whitford 1987); it may become a dominant understory plant in years with favorable moisture regimes. With increased production and density, these annual species increase the risk of fire in this community.
The ecological sites in this DRG have moderate resilience to disturbance and resistance to invasion. Primary disturbances on these sites include excessive livestock grazing, lowering of the water table, and conversion to agricultural land and urban development. Four possible stable states have been identified for this DRG.
Fire Ecology:
Fire is a rare disturbance in these plant communities, likely occurring in years with above average production. Natural fire return intervals are estimated to vary between less than 35 years up to 100 years in salt desert ecosystems with basin wildrye (Paysen et al. 2000). Historically, black greasewood-saltbush communities had sparse understories and bare soil in interspaces, making these communities somewhat resistant to fire (Young 1983, Paysen et al. 2000). They may burn only during high fire hazard conditions; for example, years with high precipitation can result in almost continuous fine fuels, increasing fire hazard (West 1994, Paysen et al. 2000).
Black greasewood may be killed by severe fires but can resprout after low to moderate severity fires (Robertson 1983, West 1994). Sheeter (1968) reported that following a Nevada wildfire, black greasewood sprouts reached approximately 2.5 feet within 3 years. Grazing and other disturbance may result in increased biomass production due to sprouting and increased seed production, also leading to greater fuel loads (Sanderson and Stutz 1994). Higher production sites would have experienced fire more frequently than lower production sites.
Shadscale is intolerant of fire and can only regenerate through seed (Zielinski 1994). Increases in the fire return interval leads to increases in the shrub component of the plant community, potentially facilitating increases in bare ground, inland salt grass and invasive weeds. Lack of fire combined with excessive herbivory decreases or eliminates the herbaceous understory, favoring black greasewood and annual species. Therefore, fire can be detrimental to these communities, especially in the presence of fire tolerant, annual non-native species.
The effect of fire on grasses relates to culm density, culm-leaf morphology, rooting characteristics, and the size of the plant. The initial condition of grasses within the site, along with seasonality and intensity of the fire, all factor into the individual species response. For most forbs and grasses the growing points are located at or below the soil surface, which provides relative protection from disturbances which decrease above ground biomass, such as grazing or fire. Thus, fire mortality is more correlated to duration and intensity of heat, which is related to culm density, culm-leaf morphology, size of plant, and abundance of old growth (Wright 1971, Young 1983). However, season and severity of the fire will influence plant response. Plant response will vary depending on post-fire soil moisture availability.
Basin wildrye is relatively resistant to fire, particularly dormant season fire, as plants sprout from surviving root crowns and rhizomes (Zschaechner 1985). Miller et al. 2013 reports fall and spring burning increased total shoot and reproductive shoot densities in the first year, although live basal areas were similar between burn and unburned plants. By year two, there was little difference between burned and control treatments.
Inland saltgrass is tolerant of fire, as its rhizomatous roots are protected beneath the soil (Monsen et al. 2004). This plant reproduces primarily through vegetative spread from rhizomesand is a poor seed producer (Monsen et al. 2004).
Alkali sacaton is tolerant of, but not resistant to fire. Recovery of alkali sacaton after fire has been reported as 2 to 4 years (Bock and Bock 1978).
Livestock/ Wildlife Grazing Interpretations:
Black greasewood is typically not considered an important browse species for wildlife and livestock. However, in a study by Smith et al. (1992), utilization of new growth on greasewood shrubs by cattle was 77 percent in summer, and greasewood was found to have the highest amounts of crude protein when compared to perennial and annual grasses. Black greasewood plants have been found to contain high amounts of sodium and potassium oxalates which are toxic to livestock and caution should be taken when grazing these communities. These shrubs can be used lightly in the spring as long as there is a substantial amount of other preferable forage available (Benson et al. 2011). Black greasewood also provides good cover for wildlife species (Benson et al. 2011).
Shadscale is a valuable browse species for a wide variety of wildlife and livestock (Blaisdell and Holmgren 1984). The spinescent growth habit of shadscale lends to its browsing tolerance with no more than 15 to 20% utilization by sheep being reported (Blaisdell and Holmgren 1984) and significantly less utilization by cattle. Increased presence of shadscale within grazed versus ungrazed areas is generally a result of the decreased competition from more heavily browsed associates (Cibils et al. 1998). Reduced competition from more palatable species in heavily grazed areas may increase shadscale germination and establishment. Chambers and Norton (1993) found shadscale establishment higher under spring than winter browsing as well as heavy compared to light browsing (p
State and transition model
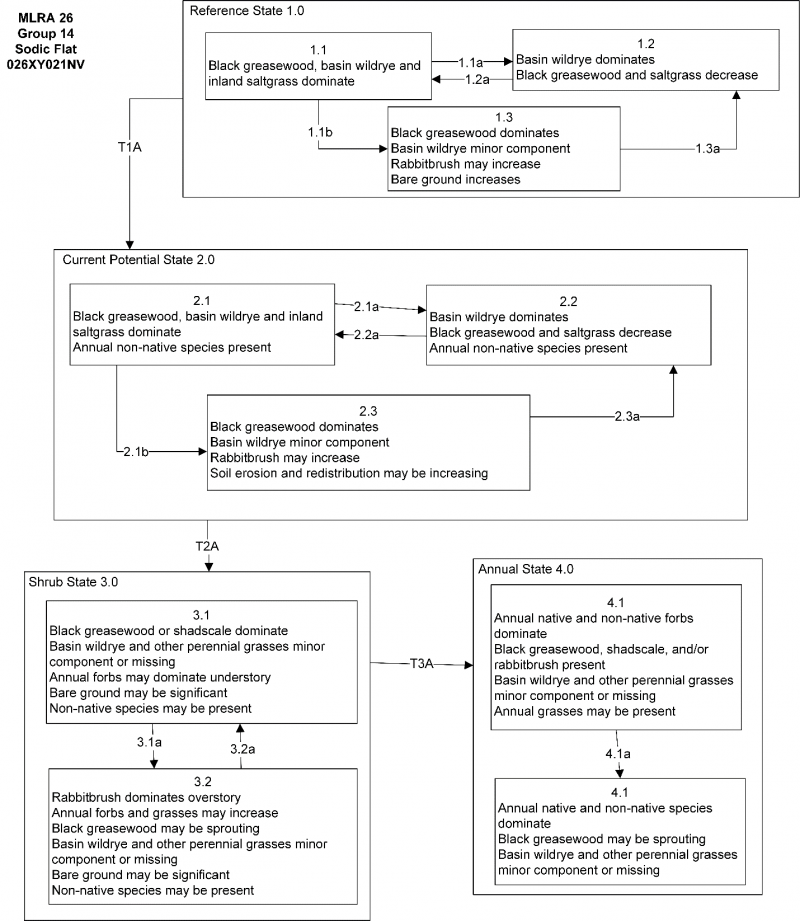

More interactive model formats are also available.
View Interactive Models
Click on state and transition labels to scroll to the respective text
Ecosystem states
State 1 submodel, plant communities
State 1
Reference State
Community 1.1
Reference Plant Community
The reference plant community is dominated by black greasewood and basin wildrye. Potential vegetative composition is about 20% grasses, 5% forbs and 75% shrubs. Approximate ground cover (basal and crown) is 20 to 25 percent.
Figure 5. Annual production by plant type (representative values) or group (midpoint values)
Table 5. Annual production by plant type
| Plant type | Low (kg/hectare) |
Representative value (kg/hectare) |
High (kg/hectare) |
|---|---|---|---|
| Shrub/Vine | 262 | 420 | 504 |
| Grass/Grasslike | 67 | 112 | 135 |
| Forb | 17 | 28 | 34 |
| Total | 346 | 560 | 673 |
Additional community tables
Table 6. Community 1.1 plant community composition
| Group | Common name | Symbol | Scientific name | Annual production (kg/hectare) | Foliar cover (%) | |
|---|---|---|---|---|---|---|
|
Grass/Grasslike
|
||||||
| 1 | Primary Perennial Grasses | 39–140 | ||||
| basin wildrye | LECI4 | Leymus cinereus | 28–84 | – | ||
| saltgrass | DISP | Distichlis spicata | 11–56 | – | ||
| 2 | Secondary Perennial Grasses | 11–45 | ||||
| squirreltail | ELEL5 | Elymus elymoides | 3–11 | – | ||
| alkali sacaton | SPAI | Sporobolus airoides | 3–11 | – | ||
|
Forb
|
||||||
| 3 | Perennial | 11–45 | ||||
| sand dropseed | SPCR | Sporobolus cryptandrus | 1–6 | – | ||
|
Shrub/Vine
|
||||||
| 4 | Primary Shrubs | 359–504 | ||||
| greasewood | SAVE4 | Sarcobatus vermiculatus | 336–420 | – | ||
| shadscale saltbush | ATCO | Atriplex confertifolia | 11–56 | – | ||
| rubber rabbitbrush | ERNA10 | Ericameria nauseosa | 11–28 | – | ||
| 5 | Secondary Shrubs | 11–45 | ||||
| Torrey's saltbush | ATTO | Atriplex torreyi | 6–11 | – | ||
| seepweed | SUAED | Suaeda | 6–11 | – | ||
| littleleaf horsebrush | TEGL | Tetradymia glabrata | 6–11 | – | ||
| shortspine horsebrush | TESP2 | Tetradymia spinosa | 6–11 | – | ||
Interpretations
Animal community
Livestock Interpretaions:
This site is suited to livestock grazing. Grazing management should be keyed to perennial bunchgrass and palatable shrub production. The early growth and abundant production of basin wildrye make it a valuable source of forage for livestock. It is important forage for cattle and is readily grazed by cattle and horses in early spring and fall. Though coarse-textured during the winter, basin wildrye may be utilized more frequently by livestock and wildlife when snow has covered low shrubs and other grasses. Saltgrass's value as forage depends primarily on the relative availability of other grasses of higher nutritional value and palatability. It can be an especially important late summer grass in arid environments after other forage grasses have deceased. Saltgrass is rated fair to good as forage species only because it stays green after most other grasses dry. Saltgrass is described as an "increaser." Black greasewood is an important winter browse plant for domestic sheep and cattle. Black greasewood may increase in response to grazing. Removal of competition can dramatically increase growth rates and total leader length of black greasewood. Shadscale is a valuable browse species, providing a source of palatable, nutritious forage for a wide variety of livestock. Shadscale provides good browse for domestic sheep. Shadscale leaves and seeds are an important component of domestic sheep and cattle winter diets. In general, livestock forage only lightly on rubber rabbitbrush during the summer, but winter use can be heavy in some locations. Fall use is variable, but flowers are often used by livestock. A few leaves and the more tender stems may also be used.
Stocking rates vary over time depending upon season of use, climate variations, site, and previous and current management goals. A safe starting stocking rate is an estimated stocking rate that is fine tuned by the client by adaptive management through the year and from year to year.
Wildlife Interpretations:
Black greasewood is an important winter browse plant for big game animals and a food source for many other wildlife species. It also receives light to moderate use by mule deer and pronghorn during spring and summer months. Black greasewood is an important source of food for many wildlife species. Shadscale is a valuable browse species, providing a source of palatable, nutritious forage for a wide variety of wildlife particularly during spring and summer before the hardening of spiny twigs. It supplies browse, seed, and cover for birds, small mammals, rabbits, deer, and pronghorn antelope. Wildlife forage only lightly on rubber rabbitbrush during the summer, but winter use can be heavy in some locations. Fall use is variable, but flowers are often used by wildlife. A few leaves and the more tender stems may also be used. The forage value of rubber rabbitbrush varies greatly among subspecies and ecotypes. Basin wildrye provides winter forage for mule deer, though use is often low compared to other native grasses. Basin wildrye provides summer forage for black-tailed jackrabbits. Because basin wildrye remains green throughout early summer, it remains available for small mammal forage for longer time than other grasses. Basin wildrye is intolerant of heavy or repeated grazing, especially if grazed before reaching maturity. Saltgrass provides cover for a variety of bird species, small mammals, and arthropods and is on occasion used as forage for several big game wildlife species.
Hydrological functions
Runoff is very high. There are no rills or pedestals on this site. Water flow patterns are rare to common dependent on location relative to major inflow areas. Water flow patterns are typically short, ending in depressional areas where water ponds. Moderately fine to fine surface textures and physical crusts result in limited infiltration rates. Concentrations of surface salts and sodium result in chemical crusts which also impede precipitation infiltration. There are "typically" no gullies associated with this site. Where this site occurs on landforms not associated with ephemeral or perennial drainageways, gullies do not occur. Where this site occurs associated with drainageways, gullies are slight to common. Occurrence of gullies is dependent on site location relative to major inflow areas. Fine litter (foliage of grasses and annual & perennial forbs) is expected to move the distance of slope length during periods of intense summer convection storms. Persistent litter (large woody material) will remain in place except during unusually deep flooding (ponding) events. This site is typically ponded for short periods in the late winter and runoff is not significant. In areas with herbaceous cover (sparse) of deep-rooted perennial herbaceous bunchgrasses (basin wildrye) and/or rhizomatous grasses (saltgrass), these plants can increase infiltration.
Recreational uses
Aesthetic value is derived from the diverse floral and faunal composition. This site is used for camping and hiking and has potential for upland and big game hunting.
Other products
The leaves, seeds and stems of black greasewood are edible. Seeds of shadscale were used by Native Americans of Arizona, Utah, and Nevada for bread and mush.
Basin wildrye was used as bedding for various Native American ceremonies, providing a cool place for dancers to stand.
Other information
Black greasewood is useful for stabilizing soil on wind-blown areas. It successfully revegetates processed oil shale and is commonly found on eroded areas and sites too saline for most plant species.
Basin wildrye is useful in mine reclamation, fire rehabilitation and stabilizing disturbed areas. Its usefulness in range seeding, however, may be limited by initially weak stand establishment.
Supporting information
Type locality
| Location 1: Douglas County, NV | |
|---|---|
| General legal description | This site also occurs in Carson City, Lyon, Mineral, Storey, and Washoe counties, Nevada. |
Other references
Alpert, P. 1990. Water sharing among ramets in a desert population of Distichlis spicata (Poaceae). American Journal of Botany 77(12):1648-1651.
Anderson, D. C., K. T. Harper, and S. R. Rushforth. 1982. Recovery of cryptogamic soil crusts from grazing on Utah winter ranges. Journal of Range Management 35:355-359.
Benson, B., D. Tilley, D. Ogle, L. St. John, S. Green, J. Briggs. 2011. Plant Guide: Black Greasewood. In: Plants database. U. S. Department of Agriculture, Natural Resources Conservation Service, Boise, ID.
Blaisdell, J. P. and R. C. Holmgren. 1984. Managing Intermountain rangelands -- salt-desert shrub ranges. Gen. Tech. Rep. INT-163. U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station, Ogden, UT. p. 52
Blauer, A. C., A. P. Plummer, E. D. McArthur, R. Stevens, and B. C. Giunta. 1976. Characteristics and hybridization of important intermountain shrubs. II. Chenopod family. USDA Forest Service Research Paper INT-177. US Department of Agriculture, Intermountain Forest and Range Experiment Station. P. 42
Bock, C. E. and J. H. Bock. 1978. Response of birds, small mammals, and vegetation to burning sacaton grasslands in southeastern Arizona. Journal of Range Management Archives 31:296-300.
Brakie, M. 2007. Plant Fact Sheet: Alkali Sacaton. In: Plants Database. U.S. Department of Agrictulture, Natural Resources Conservation Service. http://plants.usda.gov.
Bradford, J. B. and W. K. Lauenroth. 2006. Controls over invasion of Bromus tectorum: The importance of climate, soil, disturbance and seed availability. Journal of Vegetation Science 17(6): 693-704.
Branson, F. A., R. F. Miller, and I. S. McQueen. 1976. Moisture relationships in twelve northern desert shrub communities near Grand Junction, Colorado. Ecology 57:1104-1124.
Brown, R. W. 1971. Distribution of plant communities in southeastern Montana badlands. American Midland Naturalist 85:458-477.
Butler, M., F. Brummer, J. Weber, and R. Simmons. 2011. Restoring Central Oregon Rangeland from Ventenata and Medusahead to a Sustainable Bunchgrass Environment – Warm Springs and Ashwood. Central Oregon Agriculture Research and Extension Center.
Caudle, D., J. DiBenedetto, M. Karl, H. Sanchez, and C. Talbot. 2013. Interagency ecological site handbook for rangelands. Available at: http://jornada.nmsu.edu/sites/jornada.nmsu.edu/files/InteragencyEcolSiteHandbook.pdf. Accessed 4 October 2013.
Chambers, J. C. and B. E. Norton. 1993. Effects of grazing and drought on population dynamics of salt desert species on the Desert Experimental Range, Utah. Journal of Arid Environments 24:261-275.
Chambers, J., B. Bradley, C. Brown, C. D’Antonio, M. Germino, J. Grace, S. Hardegree, R. Miller, and D. Pyke. 2013. Resilience to stress and disturbance, and resistance to Bromus tectorum L. invasion in cold desert shrublands of western North America. Ecosystems 17:1-16.
Cibils, A. F., S. M. David, and D. E. McArthur. 1998. Plant-Herbivore Interactions in Atriplex: Current State of Knowledge. General Technical Report RMRS-GTR-14. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Ogden, UT. p. 31
Davies, K. W., C. S. Boyd, D. D. Johnson, A. M. Nafus, and M. D. Madsen. 2015. Success of seeding native compared with introduced perennial vegetation for revegetating medusahead-invaded sagebrush rangeland. Rangeland Ecology & Management 68(3):224-230.
Dayton, W. A. 1937. Range Plant Handbook. United States Department of Agriculture.
Devitt, D., and B. Bird. 2016. Changes in groundwater oscillations, soil water content and evapotranspiration as the water table declined in an area with deep rooted phreatophytes. Ecohydrology 9(6):1082-1093.
Eckert, R. E., Jr., A. D. Bruner, and G. J. Klomp. 1973. Productivity of tall wheatgrass and Great Basin wildrye under irrigation on a greasewood-rabbitbrush range site. Journal of Range Management 26:286-288.
Eddleman, L. E. 2002. Sarcobatus vermiculatus (Hook.) Torr.: Black greasewood. In: F. T. Bonner, [ed]. Woody plant seed manual. Department of Agriculture, Forest Service, Washington, DC.
Elmore, A. J., S. J. Manning, J. F. Mustard, and J. M. Craine. 2006. Decline in alkali meadow vegetation cover in California: the effects of groundwater extraction and drought. Journal of Applied ecology 43(4):770-779.
Fletcher, J. E. and W. P. Martin. 1948. Some effects of algae and molds in the rain-rrust of desert soils. Ecology 29:95-100.
Furbush, P. 1953. Control of medusahead on California ranges. Journal of Forestry 51(2): 118-121.
Ganskopp, D. C. 1986. Tolerances of sagebrush, rabbitbrush, and greasewood to elevated water tables. Journal of Range Management 39:334-337.
Ganskopp, D., L. Aguilera, and M. Vavra. 2007. Livestock forage conditioning among six northern Great Basin grasses. Rangeland Ecology & Management 60:71-78.
Groeneveld, D. P. 1990. Shrub rooting and water acquisition to threatened shallow groundwater habitats in the Owens Valley, California. In: D.E. McArthur, E. M. Romney, S. D. Smith, P. T. Tueller [eds.] Proceedings -- symposium on cheatgrass incasion, shrub die-off, and other aspects of shrub biology and management Gen. Tech. Rep. INT-276. U.S. Department of Agriculture, Forest Service, Intermountain Research Station, Las Vegas, NV. P. 221-237
Groeneveld, D. P. and D. E. Crowley. 1988. Root system response to flooding in three desert shrub species. Functional Ecology 2:491-497.
Gutierrez, J. R., and W. G. Whitford. 1987. Chihuahuan desert annuals: importance of water and nitrogen. Ecology 68(6):2032-2045.
Harris, G. A. 1967. Some competitive relationships between Agropyron spicatum and Bromus tectorum. Ecological Monographs 37(2): 89-111.
Hickey, W. C., Jr. and H. W. Springfield. 1966. Alkali sacaton: Its merits for forage and cover. Journal of Range Management 19:71-74.
Holmgren, R. C. and S. S. Hutchings. 1972. Salt desert shrub response to grazing use. In: C. M. McKell, J. P. Blaisdell, J. R. Goodin [eds.] Wildland shrubs- their biology and utilization. Gen. Tech. Rep. INT-1. U.S. Department of Agriculture, Intermountain Forest and Range Experiment Station. p. 153-165
Huntington, J., K. McGwire, C. Morton, K. Snyder, S. Peterson, T. Erickson, R. Niswonger, R. Carroll, G. Smith, and R. Allen. 2016. Assessing the role of climate and resource management on groundwater dependent ecosystem changes in arid environments with the Landsat archive. Remote Sensing of Environment 185:186-197.
Krall, J. L., J. R. Stroh, C. S. Cooper, and S. R. Chapman. 1971. Effect of time and extent of harvesting basin wildrye. Journal of Range Management 24:414-418.
Lei, S. A. 1999. Effects of severe drought on biodiversity and productivity in a creosote bush-blackbrush ecotone of southern Nevada. In D. E. McArthur, K. W. Ostler, C. L. Wambolt [eds.] Proceedings: shrubland ecotones. RMRS-P-11. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Ephraim, UT. p. 217-221
Mack, R. N. and D. Pyke. 1983. The demography of Bromus Tectorum: Variation in time and space. Journal of Ecology 71(1): 69-93.
Majerus, M. E. 1992. High-stature grasses for winter grazing. Journal of soil and water conservation 47:224-225.
Marcum, K. B. and D. M. Kopec. 1997. Salinity tolerance of turfgrasses and alternative species in the subfamily Chloridoideae (Poaceae). International Turfgrass Society Research Journal 8:735-742.
Meinzer, C.E. 1927. Plants as indicators of ground water. USGS Water Supply Paper 577.
Monsen, S. B., R. Stevens, and N. L. Shaw. 2004. Restoring Western Ranges and Wildlands. Gen. Tech. Rep. RMRS-GTR-136. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fort Collins, CO. p.
Mozingo, H. N. 1987. Shrubs of the Great Basin: A Natural History. In: H. N. Mozingo, [ed]. Shrubs of the Great Basin. University of Nevada Press, Reno NV. p. 67-72
Naumburg, E., R. Mata-Gonzalez, R. G. Hunter, T. Mclendon, and D. W. Martin. 2005. Phreatophytic vegetation and groundwater fluctuations: a review of current research and application of ecosystem response modeling with an emphasis on Great Basin vegetation. Environmental Management 35(6):726-740.
Parker, Karl G. 1975. Some important Utah range plants. Extension Service Bulletin EC-383. Logan, UT: Utah State University. 174 p.
Paysen, T. E., R. J. Ansley, J. K. Brown, G. J. Gottfried, S. M. Haase, M. G. Harrington, M. G. Narog, S. S. Sackett, and R. C. Wilson. 2000. Fire in western shrubland, woodland, and grassland ecosystems. In: K. C. Ryan, A. T. Jones, C. L. Koerner, K. M. Lee, [eds]. Wildland fire in ecosystems: Effects of fire on flora. Gen. Tech. Rep. RMRS-GTR-42. U.S. Department of Agriculture, Forest Service. vol 2:121-159.
Pellant, M. and C. Hall. 1994. Distribution of two exotic grasses in intermountain rangelands: status in 1992, USDA Forest Service Gen. Tech Report INT-GTR-313S: 109-112.
Ponzetti, J. M., B. McCune, and D. A. Pyke. 2007. Biotic soil crusts in relation to topography, cheatgrass and fire in the Columbia Basin, Washington. The Bryologist 110:706-722.
Robberecht, R. and G. Defossé. 1995. The relative sensitivity of two bunchgrass species to fire. International Journal of Wildland Fire 5:127-134.
Robertson, J. 1983. Greasewood (Sarcobatus vermiculatus (Hook.) Torr.). Phytologia 54:309-324.
Romo, J. T. 1984. Water relations in Artemisia tridentata subsp. wyomingensis, Sarcobatus vermiculatus, and Kochia prostrata. Thesis. Agriculture Sciences, Oregon State University, Corvallis, OR. P. 191
Roundy, B. A. 1985. Emergence and establishment of basin wildrye and tall wheatgrass in relation to moisture and salinity. Journal of Range Management 38:126-131.
Sanderson, S. C. and H. C. Stutz. 1994. Woody chenopods useful for rangeland reclamation in western North America. In: S. B. Monsen, S. G. Kitchen [eds.]Proceedings-- ecology and management of annual rangelands. Gen. Tech. Rep. INT-GTR-313. U.S. Department of Agriculture, Forest Service, Intermountain Research Station, Boise, ID. P. 374-378
Schultz, B. W. and K. W. Ostler. 1995. Effects of prolonged drought on vegetation associations in the northern Mojave Desert. In: B. A. Roundy, D. E. McArthur, J. S. Haley, D. K. Mann [eds.] Proceedings: wildland shrub and arid land restoration symposium. Gen. Tech. Rep. INT-GTR-315. U. S. Department of Agriculture, Forest Service, Intermountain Research Station, Las Vegas, NV. P. 228-235
Sheeter, G.R. 1968. Secondary succession and range improvements after wildfire in northeastern Nevada. Thesis. Reno, NV: University of Nevada. 203 p.
Sheley, R. L., E. A. Vasquez, A. Chamberlain, and B. S. Smith. 2012. Landscape-scale rehabilitation of medusahead (Taeniatherum caput-medusae)-dominated sagebrush steppe. Invasive Plant Science and Management 5(4):436-442.
Skougard, M. G., and J. D. Brotherson. 1979. Vegetational response to three environmental gradients in the salt playa near Goshen, Utah County, Utah. The Great Basin Naturalist:44-58.
Smith, M. A., J. D. Rodgers, J. L. Dodd, and Q. D. Skinner. 1992. Habitat selection by cattle along an ephemeral channel. Journal of Range Management 45:385-390.
Snyder, K. A., L. Evers, J. C. Chambers, J. Dunham, J. B. Bradford, and M. E. Loik. 2019. Effects of changing climate on the hydrological cycle in cold desert ecosystems of the Great Basin and Columbia Plateau. Rangeland Ecology & Management 72(1):1-12.
Stuart, D. M., G. E. Schuman, and A. S. Dylla. 1971. Chemical characteristics of the coppice dune soils in Paradise Valley, Nevada. Soil Sci. Soc. Am. J. 35:607-611.
Taylor, S. H., S. P. Hulme, M. Rees, B. S. Ripley, F. Ian Woodward, and C. P. Osborne. 2010. Ecophysiological traits in C3 and C4 grasses: a phylogenetically controlled screening experiment. New Phytologist 185(3):780-791.
Vest, E. D. 1962. Biotic communities in the Great Salt Lake desert. Institute of Environmental Biological Research. Ecology and Epizoology Series No. 73. Unitversity of Utah, Salt Lake City.
Weisberg, P. J., T. E. Dilts, O. W. Baughman, S. E. Meyer, E. A. Leger, K. J. Van Gunst, and L. Cleeves. 2017. Development of remote sensing indicators for mapping episodic die-off of an invasive annual grass (Bromus tectorum) from the Landsat archive. Ecological Indicators 79:173-181.
West, N. E. 1994. Effects of fire on salt-desert shrub rangelands. In: S. B. Monsen, S. G. Kitchen [eds.]Proceedings--Ecology and Management of Annual Rangelands, General Technical Report INT-313. U.S. Department of Agriculture, Forest Service, Intermountain Research Station, Boise, ID. p. 71-74
Williams, J. D. 1993. Influence of microphytic crusts on selected soil physical and hydrologic properties in the Hartnet Draw, Capital Reef National Park Utah. Dissertation. Range Science. Utah State University. Paper 2054. Available at: http://digitalcommons.usu.edu/etd/2054
Witwicki, D. L., S. M. Munson, and D. P. Thoma. 2016. Effects of climate and water balance across grasslands of varying C3 and C4 grass cover. Ecosphere 7(11).
Wright, H. A. 1971. Why squirreltail is more tolerant to burning than needle-and-thread. Journal of Range Management 24:277-284.
Young, J. A. and R. A. Evans. 1981. Germination of great basin wildrye seeds collected from native stands. Agronomy Journal 73:917-920.
Young, J. A., R. A. Evans, and P. T. Tueller. 1976. Great Basin plant communities-pristine and grazed. Holocene environmental change in the Great Basin. Nevada Archeological Survey Research Paper 6:186-215.
Young, R. P. 1983. Fire as a vegetation management tool in rangelands of the intermountain region. In S.B. Monsen, N. Shaw [eds.] Proceedings: Managing intermountain rangelands - improvement of range and wildlife habitats Gen. Tech. Rep. INT-GTR-157. U.S. Department of Agriculture, Forest Service. P. 18-31.
Zielinski, M. J. 1994. Controlling erosion on lands administered by the Bureau of Land Management, Winnemucca District, Nevada. In: S. B. Monsen, S. G. Kitchen [eds.] Proceedings - ecology and management of annual rangelands Gen. Tech. Rep. INT-GTR-313. USDA, Forest Service, Intermountain Research Station, Boise ID. p. 143-146
Zschaechner, G. A. 1985. Studying rangeland fire effects: a case study in Nevada. In: K. Sanders, J. Durham [eds.] Rangeland Fire Effects, a symposium. USDI Bureau of Land Management, Boise, Idaho. P. 66-84
Contributors
DK/FR
Tamzen Stringham
Patti Novak-Echenique
Rangeland health reference sheet
Interpreting Indicators of Rangeland Health is a qualitative assessment protocol used to determine ecosystem condition based on benchmark characteristics described in the Reference Sheet. A suite of 17 (or more) indicators are typically considered in an assessment. The ecological site(s) representative of an assessment location must be known prior to applying the protocol and must be verified based on soils and climate. Current plant community cannot be used to identify the ecological site.
| Author(s)/participant(s) | GK Brackley |
|---|---|
| Contact for lead author | State Rangeland Management Specialist |
| Date | 06/01/1979 |
| Approved by | Kendra Moseley |
| Approval date | |
| Composition (Indicators 10 and 12) based on | Annual Production |
Indicators
-
Number and extent of rills:
None -
Presence of water flow patterns:
Water flow patterns are rare to common dependent on location relative to major inflow areas. Water flow patterns are typically short, ending in depressional areas where water ponds. Moderately fine to fine surface textures and physical crusts result in limited infiltration rates. Concentrations of surface salts and sodium result in chemical crusts which also impede precipitation infiltration. -
Number and height of erosional pedestals or terracettes:
None -
Bare ground from Ecological Site Description or other studies (rock, litter, lichen, moss, plant canopy are not bare ground):
Bare Ground ± 80%; surface rock fragments less than 5%; shrub canopy less than 10%; basal area for perennial herbaceous plants ± 2%. -
Number of gullies and erosion associated with gullies:
There are "typically" no gullies associated with this site. Where this site occurs on landforms not associated with ephemeral or perennial drainageways, gullies do not occur. Where this site occurs associated with drainageways, gullies are slight to common. Occurrence of gullies is dependent on site location relative to major inflow areas. -
Extent of wind scoured, blowouts and/or depositional areas:
None -
Amount of litter movement (describe size and distance expected to travel):
Fine litter (foliage of grasses and annual & perennial forbs) is expected to move the distance of slope length during periods of intense summer convection storms. Persistent litter (large woody material) will remain in place except during unusually deep flooding (ponding) events. -
Soil surface (top few mm) resistance to erosion (stability values are averages - most sites will show a range of values):
Soil stability values will range from 1 to 4. (To be field tested.) -
Soil surface structure and SOM content (include type of structure and A-horizon color and thickness):
Structure of soil surface will be platy or massive. Soil surface colors are light grays and the soils are typified by an ochric epipedon. Organic carbon is typically less than 0.9 percent (OM values taken from lab characterization data). -
Effect of community phase composition (relative proportion of different functional groups) and spatial distribution on infiltration and runoff:
This site is typically ponded for short periods in the late winter and runoff is not significant. In areas with herbaceous cover (sparse) of deep-rooted perennial herbaceous bunchgrasses (basin wildrye) and/or rhizomatous grasses (saltgrass), these plants can increase infiltration. -
Presence and thickness of compaction layer (usually none; describe soil profile features which may be mistaken for compaction on this site):
Compacted layers are none. Platy or prismatic subsurface layers are normal for this site and are not to be interpreted as compaction. -
Functional/Structural Groups (list in order of descending dominance by above-ground annual-production or live foliar cover using symbols: >>, >, = to indicate much greater than, greater than, and equal to):
Dominant:
Reference Plant Community: Tall shrubs (black greasewood)> deep-rooted, cool season, perennial bunchgrasses = rhizomatous perennial grass. (By above ground production)Sub-dominant:
Shallow-rooted, cool season, perennial bunchgrasses > associated shrubs > deep-rooted, cool season, perennial forbs = fibrous, shallow-rooted, cool season, perennial and annual forbs. (By above ground production)Other:
Additional:
-
Amount of plant mortality and decadence (include which functional groups are expected to show mortality or decadence):
Dead branches within individual shrubs are common and standing dead shrub canopy material may be as much as 35% of total woody canopy. -
Average percent litter cover (%) and depth ( in):
Under shrubs and between plant interspaces (20-25%) and depth (± ¼ in.) -
Expected annual annual-production (this is TOTAL above-ground annual-production, not just forage annual-production):
For normal or average growing season (March thru May) ± 500lbs/ac; Winter moisture significantly affects total production. -
Potential invasive (including noxious) species (native and non-native). List species which BOTH characterize degraded states and have the potential to become a dominant or co-dominant species on the ecological site if their future establishment and growth is not actively controlled by management interventions. Species that become dominant for only one to several years (e.g., short-term response to drought or wildfire) are not invasive plants. Note that unlike other indicators, we are describing what is NOT expected in the reference state for the ecological site:
Rubber rabbitbrush, seepweed, and horsebrush are increasers. Potential invaders include cheatgrass, annual mustards, Russian thistle, halogeton and tall whitetop. -
Perennial plant reproductive capability:
All functional groups should reproduce in average (or normal) and above average growing season years.
Print Options
Sections
Font
Other
The Ecosystem Dynamics Interpretive Tool is an information system framework developed by the USDA-ARS Jornada Experimental Range, USDA Natural Resources Conservation Service, and New Mexico State University.
Click on box and path labels to scroll to the respective text.