
Natural Resources
Conservation Service
Ecological site R040XD200CA
Rarely Flooded Fans
Last updated: 3/11/2025
Accessed: 04/03/2026
General information
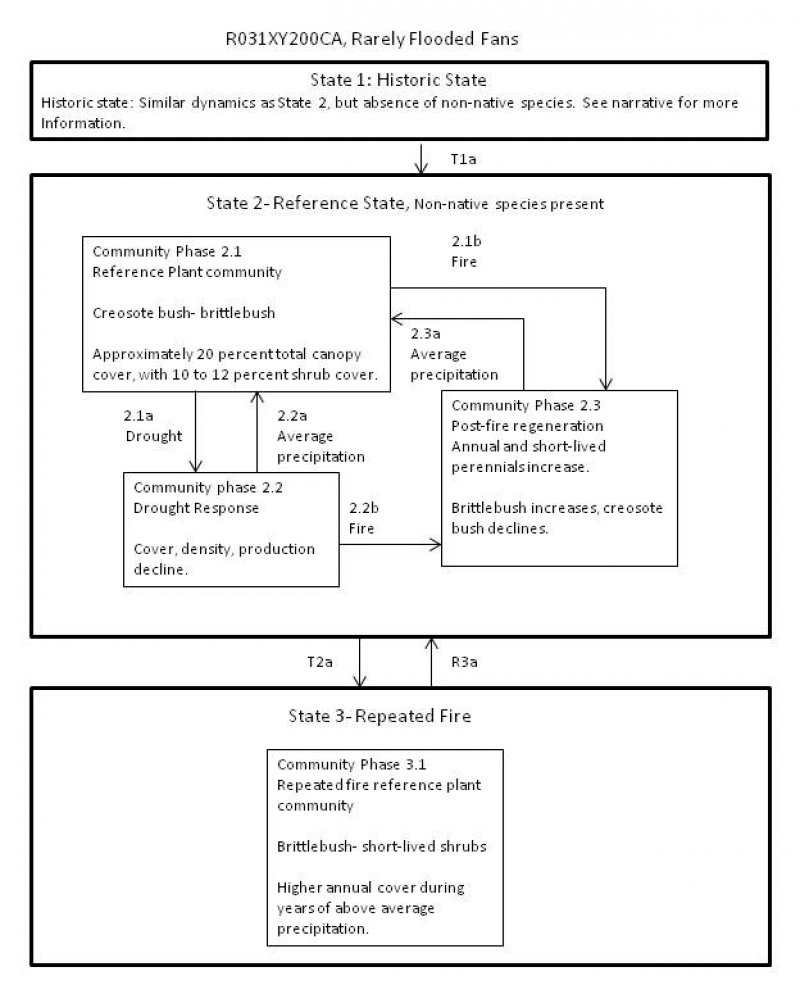
Provisional. A provisional ecological site description has undergone quality control and quality assurance review. It contains a working state and transition model and enough information to identify the ecological site.
Figure 1. Mapped extent
Areas shown in blue indicate the maximum mapped extent of this ecological site. Other ecological sites likely occur within the highlighted areas. It is also possible for this ecological site to occur outside of highlighted areas if detailed soil survey has not been completed or recently updated.
MLRA notes
Major Land Resource Area (MLRA): 040X–Sonoran Basin and Range
MLRA Statement:
Major land resource area (MLRA) 31 is the Lower Colorado Desert. This area is in the extreme southeastern part of California, in areas along the Colorado River, and in Western Arizona. The area is comprised of rough, barren, steep, and strongly dissected mountain ranges, generally northwest to southwest trending that are separated by intermontane basins. Elevation ranges from approximately 275 feet below sea level at the lowest point in the Salton Trough to 2700 feet along low northwest to southeast trending mountain ranges. The average annual precipitation is 2 to 6 inches with high temporal and spatial variability. Winter temperatures are mild, summer temperatures are hot, and seasonal and diurnal temperature fluctuations are large. Monthly minimum temperature averages range from 40 to 80 degrees F (4 to 27 degrees C). Monthly maximum temperature averages range from 65 to 110 degrees F (18 to 43 degrees C) (WRCC 2002). Temperatures are rarely below 28 degrees F, and extremely rarely fall below 24 degrees F. Precipitation is bimodal, with approximately 20 to 40 percent of annual precipitation falling between July and September. This summer rainfall, in combination with very hot temperatures and very few to no days of hard freeze are what characterize this MLRA and distinguish it from the Mojave Desert (MLRA 30).
Ecological site concept
This ecological site occurs on very rarely flooded to rarely flooded fan aprons and alluvial fans at elevations of 540 to 2730 feet. Soils are very deep and sandy. Production reference value (RV) is 218 pounds per acre and ranges from 100 to 396 pounds per acre depending on annual precipitation and annual forb production. Brittlebush and creosote bush are dominant.
Data ranges in the physiographic data, climate data, water features, and soil data sections of this Ecological Site Description are based on major components only (15 percent of map unit or greater).
Associated sites
| R040XD030CA |
Extremely Stony Fan Remnants This ecological site is on extremely stony fan remnants, near the mountain base. Teddybear cholla and a diversity of cacti and shrubs, including creosote bush and brittlebush. |
|---|---|
| R040XD034CA |
Gravelly, Braided, Ephemeral Stream This ephemeral stream is on frequently to occasionally flooded braided drainageways and channels. Burrobrush and desert lavender are dominant with a diversity of other species. |
| R040XD201CA |
Cobbly Fan Remnants This ecological site is on fan remnants with stony surfaces. Creosote bush and brittlebush are dominant, with several cacti species. |
| R040XD202CA |
Stony, Occasionally Flooded Ephemeral Stream This ephemeral stream occurs on occasionally flooded, stony, inset fans, with blue paloverde, brittlebush, creosote bush, and Schott's dalea present. |
Similar sites
| R040XD201CA |
Cobbly Fan Remnants This ecological site is also dominated by creosote bush and brittlebush, bit is it on more stable fan remnants with stony surfaces, and more cacti diversity. |
|---|---|
| R040XD005CA |
Hyper-Arid Active Alluvial Fans 2-4" p.z. This ecological site has a little more active drainage and flow patters and does not have a significant presence of brittlebush. |
Table 1. Dominant plant species
| Tree |
Not specified |
|---|---|
| Shrub |
(1) Larrea tridentata |
| Herbaceous |
Not specified |

Click on box and path labels to scroll to the respective text.