Natural Resources
Conservation Service
Ecological site R082AY366TX
Granite Hill 25-32 PZ
Last updated: 9/19/2023
Accessed: 04/03/2026
General information
Provisional. A provisional ecological site description has undergone quality control and quality assurance review. It contains a working state and transition model and enough information to identify the ecological site.
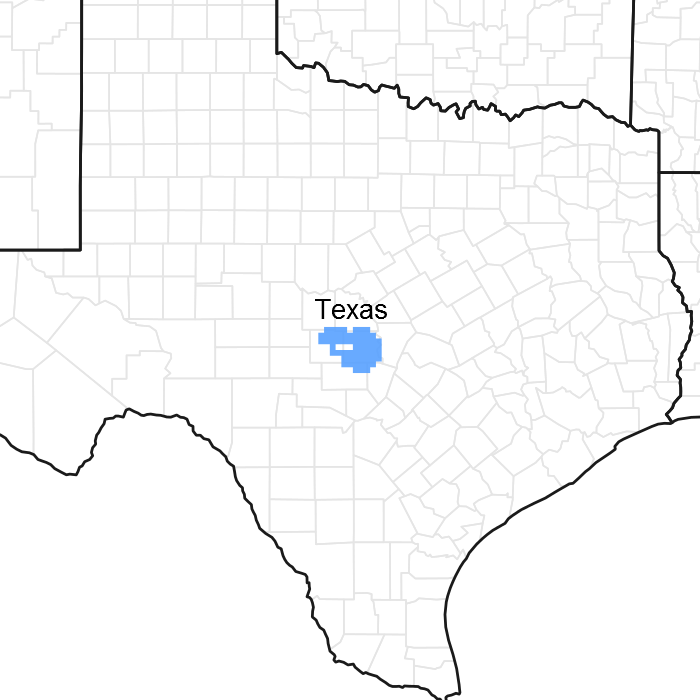
Figure 1. Mapped extent
Areas shown in blue indicate the maximum mapped extent of this ecological site. Other ecological sites likely occur within the highlighted areas. It is also possible for this ecological site to occur outside of highlighted areas if detailed soil survey has not been completed or recently updated.
MLRA notes
Major Land Resource Area (MLRA): 082A–Texas Central Basin
The 82A MLRA is underlain primarily by igneous, metamorphic, and sedimentary rocks. Igneous and metamorphic outcrops include the Valley Spring Gneiss, Packsaddle Schist, and Town Mountain Granite of Precambrian age. Sedimentary rocks include the Hickory Sandstone and Lion Mountain Sandstone of Cambrian Age and the Hessell Sand of Cretaceous age. Holocene alluvium is on flood plains.
Classification relationships
Major Land Resource Area (MLRA) and Land Resource Unit (LRU) (USDA-Natural Resources Conservation Service, 2006)
Ecological site concept
The Granite Hill ecological site is in the 25-32 inch rainfall zone. The soils are shallow over a weathered granite Cr horizon. The available water capacity is very low because of the shallow depth to parent material. Slopes range from 15 to 30 percent.
The reference vegetation consists of tall and midgrasses with an overstory savannah of oaks. Without periodic fire or brush management, woody species may increase across the site.
Associated sites
| R082AY365TX |
Granite Gravel 25-32 PZ The Granite Gravel site is deeper to parent material when compared to the Granite Hills. |
|---|---|
| R082AY368TX |
Loamy Sand 25-32 PZ The Loamy Sand site has a sandier surface and deeper soils compared to the Granite Hills. |
| R082AY373TX |
Sandy Loam 25-32 PZ The Sandy Loam site is deeper and typically not a coarse textured when compared to Granite Hills. |
| R082AY377TX |
Shallow Granite 25-32 PZ The Shallow Granite site have slopes less than 15 percent. |
Similar sites
| R082AY377TX |
Shallow Granite 25-32 PZ The Shallow Granite site occurs mostly on slopes less than 15 percent compared to Granite Hills. |
|---|
Table 1. Dominant plant species
| Tree |
(1) Quercus fusiformis |
|---|---|
| Shrub |
(1) Forestiera pubescens |
| Herbaceous |
(1) Schizachyrium scoparium |
Physiographic features
These sites occur on shallow ridges with slopes of 15 to 30 percent. The runoff on these sites is typically very high because of the slope.
Table 2. Representative physiographic features
| Landforms |
(1)
Plateau
> Ridge
(2) Hills > Hillslope |
|---|---|
| Runoff class | Very high |
| Flooding frequency | None |
| Ponding frequency | None |
| Elevation | 830 – 1,920 ft |
| Slope | 15 – 30% |
| Aspect | Aspect is not a significant factor |
Climatic features
The climate for MLRA 82A is humid subtropical and is characterized by hot summers and relatively mild winters. The average first frost should occur around November 11 and the last freeze of the season should occur around March 21.
The average relative humidity in mid-afternoon is about 50 percent. Humidity is higher at night, and the average at dawn is about 80 percent. The sun shines 70 percent of the time possible during the summer and 50 percent in winter. The prevailing wind direction is from the south.
Approximately two-thirds of the annual rainfall occurs during the April to September period. Rainfall during this period generally falls as thunderstorms, and fairly large amounts of rain may fall in localized areas for a short period of time.
Table 3. Representative climatic features
| Frost-free period (characteristic range) | 210-240 days |
|---|---|
| Freeze-free period (characteristic range) | 240-280 days |
| Precipitation total (characteristic range) | 25-32 in |
| Frost-free period (actual range) | 210-240 days |
| Freeze-free period (actual range) | 240-280 days |
| Precipitation total (actual range) | 25-32 in |
| Frost-free period (average) | 193 days |
| Freeze-free period (average) | 238 days |
| Precipitation total (average) | 28 in |
Figure 2. Monthly precipitation range
Figure 3. Monthly minimum temperature range
Figure 4. Monthly maximum temperature range
Figure 5. Monthly average minimum and maximum temperature
Figure 6. Annual precipitation pattern
Figure 7. Annual average temperature pattern
Climate stations used
-
(1) MASON [USC00415650], Mason, TX
-
(2) LLANO [USC00415272], Llano, TX
Influencing water features
These sites occur on uplands that shed water via runoff to adjacent sites. The presence of ground cover and deep-rooted grasses can help facilitate water infiltration into the soil.
Wetland description
N/A

Figure 8.
Soil features
Geology: Residuum derived from pre-Cambrian granite, gneiss and granite grus and schist.
In a representative profile for the Granite Hills ecological site, the soils are shallow over weakly or strongly cemented granite, granite grus, or gneiss. The soil profile typically has a dark brown to reddish brown coarse sandy loam with rock fragments being less than 20 percent. The permeability of the soil is moderately rapid and the permeability of the paralithic material is moderate. The water holding capacity of these soils is very low because of the depth to the root restrictive layer. These sites have a tendency to be droughty because of the very low water holding capacity. These sites occur on backslopes of ridges on dissected plateaus.
Because of the scale of mapping, there are inclusions of minor components of other soils within these mapping units. Before performing any inventories, conduct a field evaluation to ensure the soils are correct for the site.
The representative soil series associated with the Granite Hills ecological site is: Keese
Table 4. Representative soil features
| Parent material |
(1)
Residuum
–
granite
(2) Residuum – gneiss |
|---|---|
| Surface texture |
(1) Coarse sandy loam |
| Family particle size |
(1) Loamy |
| Drainage class | Well drained |
| Permeability class | Moderately rapid |
| Depth to restrictive layer | 10 – 20 in |
| Soil depth | 10 – 20 in |
| Surface fragment cover <=3" | 20% |
| Surface fragment cover >3" | 2% |
| Available water capacity (0-20in) |
0.8 – 1.8 in |
| Calcium carbonate equivalent (0-20in) |
Not specified |
| Electrical conductivity (0-20in) |
1 mmhos/cm |
| Sodium adsorption ratio (0-20in) |
2 |
| Soil reaction (1:1 water) (0-20in) |
5.6 – 6.5 |
| Subsurface fragment volume <=3" (4-20in) |
14% |
| Subsurface fragment volume >3" (4-20in) |
2% |
Ecological dynamics
The Texas Central Basin (MLRA 82A) is a unique geological region within Texas. It is composed largely of Pre-Cambrian granite, gneiss and schist (Bureau of Economic Geology 1981). Depending upon the parent material and topography, a great variety of soils have developed that vary from shallow, fissured, rocky outcrops with minimal soil development to relatively deep, well-developed soils with textures that vary from fine sandy loams to sands to gravelly clay loams to cobbly clay loams and stony clay loams (Goerdel 2000).
Precipitation patterns are highly variable. Long-term droughts, occurring three to four times per century, cause shifts in species composition by causing a die-off of seedlings, less drought-tolerant species, and some woody species. Droughts also reduce biomass production and create open space that is colonized by opportunistic species when precipitation increases. Wet periods allow little bluestem, sideoats grama, and hardwoods to increase in dominance.
The vegetation of the region developed under a humid, subtropical climate. Weather variation is great; precipitation is highly variable with seasonal, annual, and multi-year droughts (3-6 years) common as well as seasons and years with well above average precipitation; average conditions rarely exist. Typically the spring and fall are periods of highest precipitation while mid to late summer is usually a hot, droughty period. Winters are moderate with scattered precipitation sometimes in the form of short-lived snow and ice storms (Carr 1969, Bomar 1983).
Climatic variation and topographic variability interact to influence vegetation responses to disturbances such as fire and grazing. The herbaceous savannah species adapted to fire and grazing disturbances by maintaining below-ground perennating tissues. Prior to European settlement, fires would likely have been frequent (approximately every 7-12 years) (Scifres and Hamilton 1993, Frost 1998) and burned any time of year as long as there were ample fuels, dry conditions, and an ignition source.
Fire was a major influence on vegetation structure and composition prior to settlement. Lightning and Native Americans were primary ignition sources, and the latter are considered to have increased the frequency and extent of fire as their populations increased. Fires occurred at all seasons but those that occurred during the hot, dry, late-summer season following fine fuel (grass) accumulation in the spring and early summer were perhaps the most intense and had the greatest influence on the character of the vegetation. Fires were frequent, and any area may have burned once within each 7-12 year interval (Scifres and Hamilton 1993, Frost 1998). Fire generally favors the herbaceous component of the community and hinders the establishment and growth of woody species under intense hot, dry conditions. Some trees (e.g. oak species) and resprouting shrubs (e.g. mesquite) were able to escape fires, and as they matured, they became fire-resistant components of the vegetation except for infrequent stand-replacing crown fires. These woody species became effectively uncoupled from the herbaceous and shrub layer even if the herbaceous species composition was substantially altered by grazing or other factors. If, however, the oaks were killed or removed it is very difficult for them to reestablish into mature single-stemmed trees due to the resprouting nature of the tree, particularly under current land use conditions. While fire had influenced these communities for millennia, as the land was settled with homesteads and crops were established, fires were purposely prevented or stopped. Most of the remaining rangeland was overgrazed, which reduced fuel loads and hence effectively fire-proofed the plant communities from the effect of fires. This was a primary factor in the increase of woody species within the Central Basin.
Sites in reference condition are difficult to find across much of the Central Basin. While shrublands within MRLA 82 have traditionally been viewed as “degraded” relative to livestock production, it is important to recognize that they are not necessarily degraded from the ecological perspective of primary productivity, biomass accumulation, nutrient cycling, and biodiversity. The productivity of shrublands may be equal to or greater than that of the grassland they replaced. In addition, shrubs help modify soils and microclimate to increase levels of organic matter and nutrients in the upper soils horizons (Boutton et al. 2009, Boutton & Liao 2010). This nutrient enhancement by shrubs can offset grazing-induced losses of soil nutrients and contribute to enhance grass production when shrub cover is reduced. While shrub communities may have adverse impacts on grasses and grassland fauna, other plants and animals may benefit (Archer & Smeins 1991, Bestelmeyer et al. 2003). Thus, while ecosystem biodiversity certainly changes, it does not necessarily decrease with a shift from grass to woody dominance on these sites.
Soil and topographic variation interact with weather variation and land use to produce diverse plant communities across the Central Basin. Accounts of earlier explorers and settlers suggest the Central Basin was likely a mosaic of grassland, savannah, and woodlands (Foster 1917). In the reference plant community, midgrasses dominated the shortgrasses due to their ability to capture the sunlight and shade as well as being favored by the frequent fires. Plant communities vary from open grassland to savannah/parkland to shrubland/woodland to nearly closed canopy forest. Almost all sites have a two or three-layered structure of over-story trees, mid-story shrubs and a ground layer of grasses and forbs.
Historical photographs suggest the nature of the vegetation structure depending on topography, soil properties, and time since the last major disturbances (such as drought or fire). However, the occurrence of extensive grasslands and grassland fauna (pronghorn, for example) is mentioned in numerous historical accounts.
Grasses that historically dominate Central Basin sites include little bluestem (Schizachyrium scoparium), sideoats grama (Bouteloua curtipendula), meadow dropseed (Sporobolus compositus), plains lovegrass (Eragrostis intermedia), plains bristlegrass (Setaria vulpiseta), Arizona cottontop (Digitaria californica), and sand dropseed (Sporobolus cryptandrus). Locally abundant tallgrasses include Indiangrass (Sorghastrum nutans) and switchgrass (Panicum virgatum). Shortgrasses that occur in the understory of mid- and tallgrasses or on shallow soils or disturbed areas include buffalograss (Bouteloua dactyloides), common curly-mesquite (Hilaria belangeri), hairy grama (Bouteloua hirsuta), and red grama (B. trifida) (Whitehouse 1933, Riskind and Diamond 1988). The composition and productivity of grassland communities would have varied with annual rainfall, soil depth, and the extent of argillic horizon development.
Historically, overstory species composition consisted of post oak (Quercus stellata), blackjack oak (Q. marilandica), live oak (Q. virginiana), honey mesquite (Prosopis glandulosa var. glandulosa), Texas hickory (Carya texana), elm species (Ulmus spp.) and others. The shrub layer was potentially diverse with saplings of the tree layer along with whitebrush (Aloysia gratissima), lotebush (Ziziphus obtusifolia), algerita (Mahonia trifoliata), Texas persimmon (Diospyros texana), prickly pear cactus (Opuntia spp.) and others.
With the exception of Ashe juniper (Juniperus ashei), all native woody species found in the Central Basin readily resprout following fire. This trait has frustrated managers and played an important role in driving sites towards the Shrubland State. High numbers of fire sprouting shrubs make shrubland communities very resilient.
An important aspect of this site is the relationship of mature hardwood trees to each of the communities. Mature hardwoods are very resilient and remain constant whether surrounded by reference community grasslands, degraded grasslands, native-dominated shrublands, or invasive-dominated shrublands. Their presence or absence is not driven by grazing management and generally only slightly by prescribed fire. They remain relatively stable over a short management period (5-10 years) unless removed by mechanical or chemical means. Throughout this ecological site, mature oaks can occur in any of the communities if they were not historically removed. They are most likely to occur in mottes and remain relatively constant regardless of what is occurring in the rest of the community, particularly in the understory. Communities will have an absence of hardwoods if the hardwoods were harvested, burned, chained, or sprayed at some point. Once the hardwoods are removed, it is not easy to return to the historic condition due to the difficulty, expense, and time involved.
Hardwoods were frequently removed from this site during the European settlement period due to their high value for construction and firewood. Additionally, many examples exist where hardwoods were removed as part of a broad scale brush removal program. This was done with chaining, herbicides, root plowing, and other means.
Oak mottes on this site formed under different conditions than currently found. This may be due to climate shift or increased competition from aggressive shrub species. However, while reestablishment is slow, there are many examples of second-growth hardwood woodlands on this site. Hardwoods eventually reestablish when there is a lack of fire or tree clearing.
Infection of live oak by oak wilt (Ceratocystis fagacearum) has led to the death of many individuals and mottes. An increase in tree density and the grafting of roots amongst individuals has facilitated the spread of the pathogen, which is transmitted primarily through root connections (Appel 1995).
Ashe juniper, which is very abundant on the surrounding limestone derived soils of the Edwards Plateau, is relatively uncommon in the Central Basin, but it is found scattered across the Central Basin as infrequent individuals or mottes. Observation indicates that it has been increasing in population and extent within the Central Basin during the past two decades (Walter and Wyatt 1982). Juniper has the ability to take over large tracts of land as near monocultures, known as “cedar breaks.”
Even reference sites show the influence of introduced species. King Ranch bluestem (Bothriochloa ischaemum) has become almost ubiquitous, occurring on sites where it has not been seeded. It tends to replace little bluestem and can function similarly in the community as far as structure, size and soil-holding capacity. However, unlike little bluestem, King Ranch bluestem acts like an invader and moves to unoccupied areas.
The large ungulate fauna of the region prior to settlement consisted of bison (Bos bison), pronghorn antelope (Antilocarpa americana) and white-tailed deer (Odocoileus virginianus). Bison and pronghorn occasionally occurred in large numbers and may have intensively grazed the rangelands for short periods. However, they were largely migratory and free-roaming, so that when the forage became limited they moved on, often not to return for long periods. Their long-term impacts on the plant communities were considered to be relatively minor and may have had positive influences on production and diversity (Knapp et al. 1999, Fuhlendorf and Engle 2001).
While archeological evidence indicates that bison occurred in the region, there is also evidence of centuries of absence (Dillehay 1974). In addition, their numbers may have varied seasonally as herds migrated. When present, bison may have grazed certain areas heavily and then moved on. The infrequent but intense, short-duration grazing by these species suppressed woody species and invigorated herbaceous species (Eidson and Smeins 1999). After a burn, they would intensely graze the burn until no forages remained. Then, they moved off, probably not returning until the next fire cycle, which could have been 5 to 10 years. This suggests heavy short-term grazing followed by long rest periods. Activities of other native herbivores (termites, cutter ants, soil nematodes, kangaroo rats) also influenced vegetation productivity and dynamics.
Currently, white-tailed deer are primary native large herbivores. At settlement, large numbers of deer occurred, but as human populations increased (with unregulated harvest) their numbers declined substantially. Eventually, laws and restrictions on deer harvest were put in place which assisted in the recovery of the species. Females were not harvested for several decades following the implementation of hunting laws, which helped create population booms. In addition, suppression of fire favored woody plants which provided additional browse and cover for the deer. Due to their impacts on livestock production, large predators (red wolves (Canis rufus), mountain lions (Felis concolor), black bears (Ursus americanus) and eventually coyotes (Canis latrins)) were reduced in numbers or eliminated (Schmidly 2002).
The screwworm (Cochilomyia hominivorax) was essentially eradicated by the mid-1960s, and while this was immensely helpful to the livestock industry, this removed a significant control on deer populations (Teer, Thomas & Walker 1965, Bushland 1985).
Recently increased management of the deer herd, because of their economic importance through lease hunting, has decreased deer populations with the objectives of improving individual deer quality and improving habitat. High fences, controlled harvest based on numbers, sex ratios, condition, and monitoring of habitat quality have been effective in managing the deer herd on individual properties. However, across the Central Basin, excess numbers still exist which may lead to habitat degradation and significant die-offs during stress periods such as extended droughts.
The Central Basin is home to a variety of non-indigenous (exotic) ungulates, mostly introduced for hunting (Schmidly 2002). These animals are important sources of income to some landowners, but as with the white-tailed deer, their populations must be managed to prevent degradation of the habitat for themselves as well as for the diversity of native wildlife in the area. Many other species of medium and small sized mammals, birds, and insects can have significant influences on the plant communities in terms of pollination, herbivory, seed dispersal, and creation of local disturbance patches, all of which contribute to the plant species diversity.
Supplemental feeding of deer and exotics can also contribute to range degradation if it allows survival of excess numbers of animals.
Feral hogs have become well established within the Central Basin. Hogs use all of the ecological sites within MLRA 82. They cause considerable damage to soils and vegetation.
The faunal array of the Central Basin changed radically with the introduction of domestic species. Early on, wild mustangs released from early Spanish settlements roamed in large herds and had significant impacts on the vegetation. Later in the 19th century, cattle, sheep, goats, mules, and hogs were introduced. The pristine rangeland appeared to provide unlimited forage but as the ranges were fenced and overstocked they were degraded. Productivity of the rangeland began to decline, carrying capacity was reduced, and periodic die-offs of livestock occurred. Generally, the mid and taller grasses were replaced by short grasses and perennial grasses, and forbs were replaced by annuals. These changes not only reduced production but also in many instances caused permanent alteration of the ecological sites due to soil erosion, organic matter loss, compaction, moisture regime change, and other factors which altered many soil and hydrologic processes. This often precluded their recovery to pre-European conditions (Smith 1899, Smeins, Fuhlendorf and Taylor 1997). Not only did livestock overgraze the forage, but they also contributed to seed dispersal of some woody plants, particularly honey mesquite, which exacerbated its increase on the rangelands.
Historical accounts prior to the 1800s also identify grazing by herds of wild horses, followed by heavy grazing by sheep and cattle as settlement progressed. Grazing on early ranches changed natural graze-rest cycles to continuous grazing and stocking rates exceeded the carrying capacity. By the early 1800s cattle, sheep, and goat numbers appear to have been quite high in the Central Basin, resulting in heavy, year-round grazing (Lehman 1969). Sheep numbers peaked at 10.8 million head in 1943 and stood at about 1.2 million in 2000. Goat numbers in Texas around 1900 were around 100,000. They peaked in 1965 at 4.6 million and were 345,000 in 2000 (Texas Online). The Central Basin and Edwards Plateau region, because of its climate and diverse vegetation, was the mainstay of the Texas sheep and goat industry.
Today, beef cattle and horses are the primary grazers in the area. Goats used primarily for meat production are locally important, and their numbers have increased. Sheep remain a minor but still important part of livestock grazing in the Central Basin. White-tailed deer, wild turkey, bobwhite quail, and doves are major commercial wildlife species, and hunting leases are a major source of income for many landowners. While the Central Basin ecological sites have changed in many ways since settlement, opportunities exist to produce products and provide income while conserving and sustaining the long-term stability and productivity of the area.
Homesteads and communities developed along with ranching, and many ecological sites within MLRA 82 were converted to cropland for wheat (Triticum spp.), oats (Avena spp.), forage, peanuts (Arachis hypogaea), and other products needed for local consumption or for cash crops. This conversion effectively eliminated the native plant communities due to land clearing and the harvest of larger trees, used for building construction among other uses.
Over time, as many of the croplands became degraded, and along with the rangeland that had been overused, introduced forages were brought in to assist with soil and water conservation and to increase productivity. Coastal bermudagrass (Cynodon dactylon), Kleingrass (Panicum coloratum), Wilman lovegrass (Eragrostis superba), and King Ranch bluestem were widely planted on many acres of old cropland and in areas with deeper soils. The latter, while effective as a soil stabilizer, has become invasive in many areas, including sites with shallower soils. It is difficult to control.
In the 1940s, mechanical and herbicide treatments began to replace fire as a control of increasing density of woody plants on the rangeland. This activity was common practice for several decades until the 1980s, when these treatments became less cost-effective. It was clear that brush management practices were treating symptoms rather than underlying problems in addition to their undesirable environmental and wildlife consequences. Sites cleared of brush regenerated rapidly and often formed thickets that were denser and of lower diversity than the original stands. This realization coupled with the fact that brush management treatments were typically expensive and short-lived, lead to the development of Integrated Brush Management Systems (Scifres et al. 1985). This approach takes a holistic, large-scale, long-term, socioeconomic, ecosystem-based approach to brush management and recognizes multiple-use options for rangeland resources including alternate classes of livestock, lease hunting, exotic game ranching, carbon credits and ecotourism.
Grazing and fire are two factors that critically influence the relative abundance of grasses and woody plants through time. The resulting reduction in abundance of late seral grasses lead to a decline in soil organic matter, a reduction in fire frequency/intensity (due to lack of fine fuels), and a shift in dominance from midgrasses (little bluestem and sideoats grama) to shortgrasses (hooded windmillgrass (Chloris cucullata) and buffalograss) and forbs {Mexican sagewort (Artemisia ludoviciana ssp. mexicana) and croton (Croton spp.)}. These changes would have favored woody plants, most of which are unpalatable to livestock, and enabled them to establish and maintain dominance.
Mesquite, whitebrush, juniper, lotebush, algerita, persimmon, prickly pear, and lime pricklyash (Zanthoxylum fagar) now dominate much of the Central Basin. These woody plants are not ‘new arrivals’ but rather, are native to the region and have increased in size and abundance within their historic ranges. Factors promoting their increase in abundance since European settlement are the subject of active debate. Such factors may involve an interactive combination of changes in climate, intensification of grazing; follow up brush management and reductions in fire frequency/intensity accompanied by increases in atmospheric CO2 concentrations and nitrogen deposition since the industrial revolution (Archer 1994).
A State and Transition Model diagram for the Granite Hills is depicted in Figure 1. Thorough descriptions of each state, transition, plant community, and pathway follow the model. Experts base this model on available experimental research, field observations, professional consensus, and interpretations. It is likely to change as knowledge increases.
Plant communities will differ across the MLRA due to the naturally occurring variability in weather, soils, and aspect. The Savannah State is the reference state for this site. It is not necessarily the management goal but can be. Other vegetative states may be desired plant communities as long as the Range Health assessments are in the moderate and above category. The biological processes on this site are complex. Therefore, representative values are presented in a land management context. The species lists are representative and are not botanical descriptions of all species occurring, or potentially occurring, on this site. They are not intended to cover every situation or the full range of conditions, species, and responses for the site.
Both percent species composition by weight and percent canopy cover are used in this ESD. Most observers find it easier to visualize or estimate percent canopy for woody species (trees and shrubs). Canopy cover drives the transitions between communities and states because of the influence of shade and interception of rainfall. Species composition by dry weight is used for describing the herbaceous community and the community as a whole. Woody species are included in species composition for the site. Calculating similarity index requires the use of species composition by dry weight.
The following diagram suggests some pathways that the vegetation on this site might take. There may be other states not shown in the diagram. This information is intended to show what might happen in a given set of circumstances; it does not mean that this would happen the same way in every instance. Local professional guidance should always be sought before pursuing a treatment scenario.
State and transition model
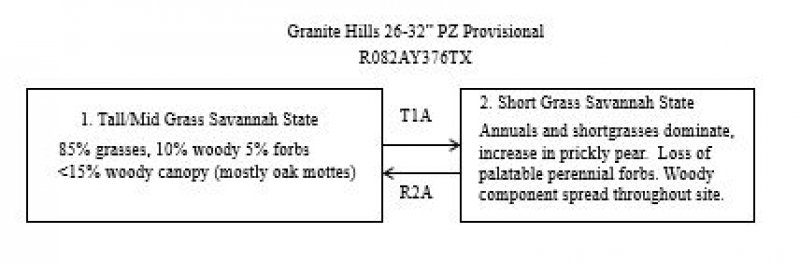
Figure 9. State and Transitioin Model diagram
More interactive model formats are also available.
View Interactive Models
More interactive model formats are also available.
View Interactive Models
Click on state and transition labels to scroll to the respective text
Ecosystem states
| T1A | - | Absence of disturbance, natural regeneration over time, and prolonged excessive grazing pressure |
|---|---|---|
| R2A | - | Removal of woody canopy and reintroduction of historic disturbance return intervals |
State 1 submodel, plant communities
State 1
Tall/Mid Grass Savannah State
This is the reference state for the Granite Hills ecological site. The information is derived from a range site description approved April 16, 1985. The plant community is a post oak and live oak savannah. The understory is dominated by tall and mid grasses such as little bluestem, sand lovegrass, sideoats grama, and green sprangletop. The oak overstory shades about 20 percent of the ground. This site supports an abundance of forbs with some woody shrubs and vines. The north slopes grow a higher percent of little bluestem and oaks than south slopes. The vegetation grows in the soil between the rock outcrops. In the Tall/Mid Grass Savannah State, the grasses soak up much of the sunlight and their fair share of the soil moisture as long as the woody canopy remains less than 15 percent. If no brush management is done, or if fire is not utilized, the woody canopy will continue to increase until a threshold is crossed at about 20 percent canopy. Once this happens, it will take mechanical or chemical intervention to restore the Tall/Mid Grass Savannah State community.
Dominant plant species
-
Texas live oak (Quercus fusiformis), tree
-
little bluestem (Schizachyrium scoparium), grass
-
sideoats grama (Bouteloua curtipendula), grass
Community 1.1
Tall/Mid Grass Savannah
The site has 85 percent grasses, 10 percent woody, 5 percent forbs, less than 15 percent woody canopy, mostly in oak mottes.
Figure 10. Annual production by plant type (representative values) or group (midpoint values)
Table 5. Annual production by plant type
| Plant type | Low (lb/acre) |
Representative value (lb/acre) |
High (lb/acre) |
|---|---|---|---|
| Grass/Grasslike | 850 | 1275 | 1700 |
| Tree | 100 | 150 | 200 |
| Forb | 50 | 75 | 100 |
| Shrub/Vine | 10 | 30 | 50 |
| Total | 1010 | 1530 | 2050 |
State 2
Short Grass Savannah State
As retrogression occurs, juniper, Texas persimmon, whitebrush, and mesquite may form a dense canopy. Some common increasers or invaders on the are signalgrass, basin sneezeweed, pricklypear, tasajillo, mesquite and certain annuals. Unless removed by prior mechanical or chemical treatment, the oaks and trees remain on the site at about a 15 percent canopy because of the relatively small amounts of soil on the site to support trees. The understory shrubs begin to increase in canopy and can be more than 15 percent. Once this condition exists, completion from woody species and an overstory reduces the herbaceous component of the plant community. The sun’s energy is captured by the woody species and most of the available soil moisture is utilized by the woody species.
Dominant plant species
-
juniper (Juniperus), tree
-
mesquite (Prosopis), tree
-
Texas persimmon (Diospyros texana), shrub
Transition T1A
State 1 to 2
Transition T1A is a result of the loss of sunlight energy capture by tall and midgrasses that is captured by woody species. A lack of mineral cycling contributes to this transition. This can occur as a result of abusive grazing, lack of fire, and a lack of brush management. Droughts will hasten the process.
Restoration pathway R2A
State 2 to 1
Restoration pathway R2A is a result of intervention with brush management and prescribed grazing. Prescribed grazing will build fuel load, to allow the use of prescribed fire as a tool.
Conservation practices
| Brush Management | |
|---|---|
| Prescribed Burning | |
| Prescribed Grazing |
Additional community tables
Table 6. Community 1.1 plant community composition
| Group | Common name | Symbol | Scientific name | Annual production (lb/acre) | Foliar cover (%) | |
|---|---|---|---|---|---|---|
|
Grass/Grasslike
|
||||||
| 1 | Tall/ Mid Grasses | 755–1520 | ||||
| little bluestem | SCSC | Schizachyrium scoparium | 455–920 | – | ||
| sideoats grama | BOCU | Bouteloua curtipendula | 100–200 | – | ||
| Indiangrass | SONU2 | Sorghastrum nutans | 50–100 | – | ||
| purpletop tridens | TRFL2 | Tridens flavus | 30–60 | – | ||
| sand dropseed | SPCR | Sporobolus cryptandrus | 20–40 | – | ||
| plains bristlegrass | SEVU2 | Setaria vulpiseta | 20–40 | – | ||
| plains lovegrass | ERIN | Eragrostis intermedia | 20–40 | – | ||
| sand lovegrass | ERTR3 | Eragrostis trichodes | 20–40 | – | ||
| tanglehead | HECO10 | Heteropogon contortus | 20–40 | – | ||
| green sprangletop | LEDU | Leptochloa dubia | 10–20 | – | ||
| Arizona cottontop | DICA8 | Digitaria californica | 10–20 | – | ||
| 2 | perennial cool season | 50–100 | ||||
| sedge | CAREX | Carex | 25–75 | – | ||
| wildrye | ELYMU | Elymus | 25–75 | – | ||
| 3 | Short grasses | 50–100 | ||||
| silver bluestem | BOSA | Bothriochloa saccharoides | 25–75 | – | ||
| hooded windmill grass | CHCU2 | Chloris cucullata | 25–50 | – | ||
| tumble windmill grass | CHVE2 | Chloris verticillata | 25–50 | – | ||
| roughhair rosette grass | DISTL | Dichanthelium strigosum var. leucoblepharis | 25–50 | – | ||
| hairy grama | BOHI2 | Bouteloua hirsuta | 25–50 | – | ||
|
Tree
|
||||||
| 4 | Trees | 80–160 | ||||
| black hickory | CATE9 | Carya texana | 10–20 | – | ||
| sugarberry | CELA | Celtis laevigata | 10–20 | – | ||
| Texas live oak | QUFU | Quercus fusiformis | 10–20 | – | ||
| blackjack oak | QUMA3 | Quercus marilandica | 10–20 | – | ||
| post oak | QUST | Quercus stellata | 10–20 | – | ||
| saw greenbrier | SMBO2 | Smilax bona-nox | 10–20 | – | ||
| yucca | YUCCA | Yucca | 10–20 | – | ||
|
Shrub/Vine
|
||||||
| 5 | shrubs | 20–40 | ||||
| Texas kidneywood | EYTE | Eysenhardtia texana | 5–10 | – | ||
| stretchberry | FOPU2 | Forestiera pubescens | 5–10 | – | ||
| western white honeysuckle | LOAL | Lonicera albiflora | 5–10 | – | ||
| fragrant mimosa | MIBO2 | Mimosa borealis | 5–10 | – | ||
| Texas almond | PRMI2 | Prunus minutiflora | 5–10 | – | ||
| winged sumac | RHCO | Rhus copallinum | 5–10 | – | ||
| gum bully | SILAL3 | Sideroxylon lanuginosum ssp. lanuginosum | 5–10 | – | ||
|
Forb
|
||||||
| 6 | Forbs | 50–100 | ||||
| white sagebrush | ARLU | Artemisia ludoviciana | 25–60 | – | ||
| Texas lupine | LUTE | Lupinus texensis | 0–50 | – | ||
| awnless bushsunflower | SICA7 | Simsia calva | 20–50 | – | ||
| Cuman ragweed | AMPS | Ambrosia psilostachya | 25–50 | – | ||
| littleleaf sensitive-briar | MIMI22 | Mimosa microphylla | 20–40 | – | ||
| Engelmann's daisy | ENPE4 | Engelmannia peristenia | 20–40 | – | ||
| ratany | KRAME | Krameria | 10–30 | – | ||
| amberique-bean | STHE9 | Strophostyles helvola | 10–30 | – | ||
| fringeleaf wild petunia | RUHU | Ruellia humilis | 10–25 | – | ||
| croton | CROTO | Croton | 0–25 | – | ||
Interpretations
Animal community
Game birds, songbirds, and birds of prey were indigenous or frequent users, and most are still plentiful. Quail and doves frequent this site depending upon the vegetative community. Small mammals that use the site include armadillos, opossum, raccoons, rodents, jackrabbits, cottontail rabbits, and skunks. Its use by deer is limited by browse and cover in reference condition. As ecological state shifts and woody plants increase and invade, it becomes more habitable for deer. Deer prefer many of the forbs and legumes that grow on the site.
Feral hogs (Sus scrofa) can be found on most Ecological Sites in Texas. Damage is caused by feral hogs each year including, crop damage by rutting up crops, destroyed fences, livestock watering areas, and predation on native wildlife, domestic livestock (small calves, goats, and sheep) and ground-nesting birds. Feral hogs have no natural predators other than humans, thus allowing their population to grow to high numbers (Cearley 2009 & Mapston 2004). Feral hogs have naturalized to rangelands across the state.
Predators including bobcats, coyotes, foxes, and mountain lions can also be found on the site.
Hydrological functions
Small rains can be utilized because runoff from rocks is absorbed in the soil. Large rains generally produce runoff. The presence of good ground cover and deep-rooted grasses can help facilitate infiltration and reduce sediment loss.
Supporting information
Inventory data references
USDA Range Site Description. January 17, 1985, Granite Hills PE 38 – 49, Central Basin, Burnet, Johnson City, Fredericksburg, Llano, Mason Counties.
Other references
AgriLife. Wildlife. "Managing Feral Hogs Not a One-shot Endeavor." Press release. AgNews. 01 Jan. 2009. Texas Cooperative Extension. 23 Apr. 2009 (http://agnews.tamu.edu/showstory.php?id=903).
Appel, D. N. 1995. The Oak Wilt Enigma: Perspective from the Texas Epidemic. Ann. Rev. Phytopathol. 33:103-118.
Archer, S. 1994. Woody plant encroachment into southwestern grasslands and savannas: rates, patterns and proximate causes. In: Ecological implications of livestock herbivory in the West, pp. 13-68. Edited by M. Vavra, W. Laycock, R. Pieper. Society for Range Management Publication, Denver, CO.
Archer, S. and F. Smeins. 1991. Ecosystem-Level Processes. Pp. 109-139, In Grazing Management: An Ecological Perspective. Edited by R.K. Heitschmidt and J.W. Stuth. Timber Press, Inc., Portland. 259p.
Bestelmeyer, B.T., J.R. Brown, K.M. Havstad, R. Alexander, G. Chavez and J.E. Hedrick. 2003. Development and Use of State-and-Transition Models for Rangelands. J. Range Manage. 56: 114-126.
Bomar, G.W. 1983. Texas Weather. Univ. Tex. Press, Austin. 265p.
Brown, J.R. and S. Archer. 1999. Shrub invasion of grassland: recruitment is continuous and not regulated by herbaceous biomass or density. Ecology 80(7): 2385-2396.
Bureau of Economic Geology. 1981. Geologic Atlas of Texas, Llano Sheet. Bur. Econ. Geol., Univ. Tex. Austin.
Bushland, R.C. 1985. Eradication program in the southwestern United States. Symposium on eradication of the screwworm from the United States and Mexico. Misc. Pub. Entomol. Soc. Am., 62:12-15.
Carr, J.T. 1969. The Climate and Physiography of Texas. Tex. Water Devel. Bd. Rep. No. 53. 27p.
Eidson, J.A. and F.E. Smeins. 1999. Texas blackland prairies. 305–307. in Terrestrial ecoregions of North America: a conservation assessment. Ricketts, T., E. Dinerstein, and D. Olson. editors. Island Press. Washington, D.C.
Everitt, J.H., D.L. Drawe, and R.I. Lonard. 1999. Field Guide to the Broad-Leaved Herbaceous Plants of South Texas. Lubbock, Texas: Texas Tech University Press.
Everitt, J.H., D.L. Drawe, and R.I. Lonard. 2002. Trees, Shrubs, and Cacti of South Texas. Lubbock, Texas: Texas Tech University Press.
Foster, J.H. 1917. Pre-settlement fire frequency regions of the United States: a first approximation. Tall Timbers Fire Ecology Conference Proceedings No. 20.
Foster, J.H. 1917. The Spread of Timbered Areas in Central Texas. J. For. 15:442-445.
Frost, C. C. 1995. Presettlement fire regimes in southeastern marshes, peatlands, and swamps. In: Proceedings, 19th Tall Timbers fire ecology conference. Tallahassee, FL: Tall Timbers Research Station pp. 39-60.
Frost, C. C. 1998. Pre-settlement fire frequency regions of the United States: A first approximation. Tall Timbers Fire Ecology Conference Proceedings No. 20.
Fuhlendorf, S. D. and D. M. Engle. 2001. Restoring Heterogeneity on Rangelands: Ecosystem Management Based on Evolutionary Grazing Patterns. Bioscience. 51:625-632.
Fulbright, T. E., J. A. Ortega-Santos, A. Lozano-Cavazos, and L. E. Ramírez-Yánez. 2006. Establishing vegetation on migrating inland sand dunes in Texas. Rangeland Ecology and Management 59:549-556.
Goerdel, A.R. 2000. Soil Survey of Llano County. USDA, Natural Resources Conservation Service, Washington, D.C.
Gould, F.W. 1975. The Grasses of Texas. Texas A&M University Press, College Station, TX. 653p.
Grace, J. B., L. K. Allain, H. Q. Baldwin, A. G. Billock, W. R. Eddleman, A. M. Given, C. W. Jeske, and R. Moss. 2005. Effects of prescribed fire in the coastal prairies of Texas. USGS Open File Report 2005-1287.
Hanselka, W., R. Lyons, and M. Moseley. 2009. Grazing Land Stewardship – A Manual for Texas Landowners. Texas AgriLife Communications, HTTP://AGRILIFEBOOKSTORE.ORG.
Hart, C. R. t. Garland, A. C. Barr, B. B. Carpenter and J. C. Reagor. 2003. Toxic Plants of Texas. Texas Cooperative Extension Bulletin B-6103 11-03.
Knapp, A.K., et al. 1999. The Keystone Role of Bison in North American Tallgrass Prairie. Bioscience 49: 39-50.
Kneuper, C.L., C.B. Scott, and W.E. Pinchak. 2003. Consumption and Dispersion of Mesquite Seeds by Ruminants. Journal of Range Management. 56:255-259.
Kramp, B, R, Ansley, and D. Jones. 1999. The effect of prescribed fire on mesquite seedlings. Vernon Center Technical report.
Mann, C. 2004. 1491. New Revelations of the Americas before Columbus.
Mapston, Mark E. Feral Hogs in Texas. Rep. Texas Cooperative Extension. 23 Apr. 2009 (http://icwdm.org/Publications/pdf/Feral%20Pig/Txferalhogs.pdf)=
Riskind, D.H. and D.D. Diamond. 1988. An Introduction to Environment and Vegetation. Pp. 1-15, In Edwards Plateau Vegetation: Plant Ecological Studies in Central Texas. Edited by B.B. Amos and F.R. Gehlbach. Baylor University Press, Waco, TX.
Scifres, C.J. and W.T. Hamilton. 1993. Prescribed Burning for Brush Management: The South Texas Example. Texas A&M University Press, College Station, TX. 245 p.
Scifres, C.J., H.T. Hamilton, J.R. Conner, J.M Inglis, G.A. Rasumssen, R.P. Smith, J.W. Stuth, T.G. Welch (eds.) 1985. Integrated brush management Systems for South Texas: Development and implementation. Tex. Ag. Exp. Stat. B-1493. 71 p.
Smeins, F., S. Fuhlendorf, and C. Taylor, Jr. 1997. Environmental and Land Use Changes: A Long Term Perspective. Chapter 1 in: Juniper Symposium 1997, pp. 1-21. Texas Agricultural Experiment Station.
Smith, J.G. 1899. Grazing Problems in the Southwest and How To Meet Them. U.S. Dep. Agr. Div. Agron. Bull. No. 16. 47p.
Stringham, T.K., W.C. Krueger, and P.L. Shaver. 2001. State and transition modeling: an ecological process approach. J. Range Management. 56(2):106-113.
Teer, J.G., J.W. Thomas and E.A. Walker. 1965. Ecology and Management of White-tailed Deer in the Llano Basin of Texas. Wildlife Monographs 10: 1-62.
Texas A&M Research and Extension Center. 2000. Native Plants of South Texas (http://uvalde.tamu.edu/herbarium/index.html).
Texas Agriculture Experiment Station. 2007. Benny Simpson’s Texas Native Trees (http://aggie-horticulture.tamu.edu/ornamentals/natives/).
Texas Online. http://www.tshaonline.org/handbook/online/articles/asw02
Texas Parks and Wildlife Dept. 2007. List of White-tailed Deer Browse and Ratings. District 8.
Thurow, T.L. 1991. Hydrology and Erosion. Chapter 6 in: Grazing Management: An Ecological Perspective. Edited by R.K. Heitschmidt and J.W. Stuth. Timber Press, Portland, OR.
TR 1737-15 (1998) Riparian Area Management – a User’s Guide to Assessing Proper Functioning Condition and the Supporting Science for Lotic Areas. Bureau of Land Management, US Forest Service, Natural Resources Conservation Service.
USDA, NRCS. 1997. National Range and Pasture Handbook.
USDA, NRCS. 2007. The PLANTS Database (http://plants.usda.gov). National Plant Data Center, Baton Rouge, LA 70874-4490 USA.
USDA, NRCS (Formerly Soil Conservation Service) Range Site Description. January 17, 1985, Granite Hills PE 38 – 49, Central Basin, Burnet, Johnson City, Fredericksburg, Llano, Mason Counties. Range site description approved by Herb Senne, State Rangeland Management Specialist, 4-11-85, Lester C. Brockman, TSS – Soil Scientist, 4-16-85, and Jackie Elrod, Area Conservationist, 4-26-85.
USDA/NRCS Soil Survey Manuals for appropriate counties within MLRA 86A.
Vines, R.A. 1977. Trees of Eastern Texas. University of Texas Press, Austin, TX. 538 p.
Vines, R.A. 1984. Trees of Central Texas. University of Texas Press, Austin, TX.
Wade, D. D., B. L. Brock, P. H. Brose, J. B. Grace, G. A. Hoch, and W. A. Patterson III. 2000. Fire in Eastern ecosystems. In: Brown, J.K., and J. Kaplers, eds. Wildland fire in ecosystems: effects of fire on flora. Gen. Tech. Rep. RMRS-GTR-42-vol. 2. Ogden, UT: United States Department of Agriculture, Forest Service, Rocky Mountain Research Station 257 p.
Weniger, D. 1984. The Explorers’ Texas: The Land and Waters. Eakin Press, Austin. 224 p.
Whitehouse, E. 1933. Plant Succession on Central Texas Granite. Ecol. 14: 391-404.
Wright, H.A. and A.W. Bailey. 1982. Fire Ecology: United States and Southern Canada. John Wiley & Sons, Inc.
Wright, B.D., R.K. Lyons, J.C. Cathey, and S. Cooper. 2002. White-tailed Deer Browse Preferences for South Texas and the Edwards Plateau. Texas Cooperative Extension Bulletin B-6130.
Contributors
Mark Moseley
Edits by Travis Waiser, MLRA Leader, NRCS, Kerrville, TX
Approval
Bryan Christensen, 9/19/2023
Acknowledgments
Site Development and Testing Plan:
Future work, as described in a Project Plan, to validate the information in this Provisional Ecological Site Description is needed. This will include field activities to collect low, medium and high intensity sampling, soil correlations, and analysis of that data. Annual field reviews should be done by soil scientists and vegetation specialists. A final field review, peer review, quality control, and quality assurance reviews of the ESD will be needed to produce the final document. Annual reviews of the Project Plan are to be conducted by the Ecological Site Technical Team.
QC/QA completed by:
Bryan Christensen, SRESS, NRCS, Temple, TX
Erin Hourihan, ESDQS, NRCS, Temple, TX
Rangeland health reference sheet
Interpreting Indicators of Rangeland Health is a qualitative assessment protocol used to determine ecosystem condition based on benchmark characteristics described in the Reference Sheet. A suite of 17 (or more) indicators are typically considered in an assessment. The ecological site(s) representative of an assessment location must be known prior to applying the protocol and must be verified based on soils and climate. Current plant community cannot be used to identify the ecological site.
| Author(s)/participant(s) | |
|---|---|
| Contact for lead author | |
| Date | 09/20/2023 |
| Approved by | Bryan Christensen |
| Approval date | |
| Composition (Indicators 10 and 12) based on | Annual Production |
Indicators
-
Number and extent of rills:
-
Presence of water flow patterns:
-
Number and height of erosional pedestals or terracettes:
-
Bare ground from Ecological Site Description or other studies (rock, litter, lichen, moss, plant canopy are not bare ground):
-
Number of gullies and erosion associated with gullies:
-
Extent of wind scoured, blowouts and/or depositional areas:
-
Amount of litter movement (describe size and distance expected to travel):
-
Soil surface (top few mm) resistance to erosion (stability values are averages - most sites will show a range of values):
-
Soil surface structure and SOM content (include type of structure and A-horizon color and thickness):
-
Effect of community phase composition (relative proportion of different functional groups) and spatial distribution on infiltration and runoff:
-
Presence and thickness of compaction layer (usually none; describe soil profile features which may be mistaken for compaction on this site):
-
Functional/Structural Groups (list in order of descending dominance by above-ground annual-production or live foliar cover using symbols: >>, >, = to indicate much greater than, greater than, and equal to):
Dominant:
Sub-dominant:
Other:
Additional:
-
Amount of plant mortality and decadence (include which functional groups are expected to show mortality or decadence):
-
Average percent litter cover (%) and depth ( in):
-
Expected annual annual-production (this is TOTAL above-ground annual-production, not just forage annual-production):
-
Potential invasive (including noxious) species (native and non-native). List species which BOTH characterize degraded states and have the potential to become a dominant or co-dominant species on the ecological site if their future establishment and growth is not actively controlled by management interventions. Species that become dominant for only one to several years (e.g., short-term response to drought or wildfire) are not invasive plants. Note that unlike other indicators, we are describing what is NOT expected in the reference state for the ecological site:
-
Perennial plant reproductive capability:
Print Options
Sections
Font
Other
The Ecosystem Dynamics Interpretive Tool is an information system framework developed by the USDA-ARS Jornada Experimental Range, USDA Natural Resources Conservation Service, and New Mexico State University.
Click on box and path labels to scroll to the respective text.
Ecosystem states
| T1A | - | Absence of disturbance, natural regeneration over time, and prolonged excessive grazing pressure |
|---|---|---|
| R2A | - | Removal of woody canopy and reintroduction of historic disturbance return intervals |