Natural Resources
Conservation Service
Ecological site R026XY009NV
MAHOGANY SAVANNA
Last updated: 4/10/2024
Accessed: 04/04/2026
General information
Provisional. A provisional ecological site description has undergone quality control and quality assurance review. It contains a working state and transition model and enough information to identify the ecological site.
MLRA notes
Major Land Resource Area (MLRA): 026X–Carson Basin and Mountains
The area lies within western Nevada and eastern California, with about 69 percent being within Nevada, and 31 percent being within California. Almost all this area is in the Great Basin Section of the Basin and Range Province of the Intermontane Plateaus. Isolated north-south trending mountain ranges are separated by aggraded desert plains. The mountains are uplifted fault blocks with steep side slopes. Most of the valleys are drained by three major rivers flowing east across this MLRA. A narrow strip along the western border of the area is in the Sierra Nevada Section of the Cascade-Sierra Mountains Province of the Pacific Mountain System. The Sierra Nevada Mountains are primarily a large fault block that has been uplifted with a dominant tilt to the west. This structure leaves an impressive wall of mountains directly west of this area. This helps create a rain shadow affect to MLRA 26. Parts of this eastern face, but mostly just the foothills, mark the western boundary of this area. Elevations range from about 3,806 feet (1,160 meters) on the west shore of Pyramid Lake to 11,653 feet (3,552 meters) on the summit of Mount Patterson in the Sweetwater Mountains.
Valley areas are dominantly composed of Quaternary alluvial deposits with Quaternary playa or alluvial flat deposits often occupying the lowest valley bottoms in the internally drained valleys, and river deposited alluvium being dominant in externally drained valleys. Hills and mountains are dominantly Tertiary andesitic flows, breccias, ash flow tuffs, rhyolite tuffs or granodioritic rocks. Quaternary basalt flows are present in lesser amounts, and Jurassic and Triassic limestone and shale, and Precambrian limestone and dolomite are also present in very limited amounts. Also of limited extent are glacial till deposits along the east flank of the Sierra Nevada Mountains, the result of alpine glaciation.
The average annual precipitation in this area is 5 to 36 inches (125 to 915 millimeters), increasing with elevation. Most of the rainfall occurs as high-intensity, convective storms in spring and autumn. Precipitation is mostly snow in winter. Summers are dry. The average annual temperature is 37 to 54 degrees F (3 to 12 degrees C). The freeze-free period averages 115 days and ranges from 40 to 195 days, decreasing in length with elevation.
The dominant soil orders in this MLRA are Aridisols and Mollisols. The soils in the area dominantly have a mesic soil temperature regime, an aridic or xeric soil moisture regime, and mixed or smectitic mineralogy. They generally are well drained, are clayey or loamy and commonly skeletal, and are very shallow to moderately deep.
This area supports shrub-grass vegetation characterized by big sagebrush. Low sagebrush and Lahontan sagebrush occur on some soils. Antelope bitterbrush, squirreltail, desert needlegrass, Thurber needlegrass, and Indian ricegrass are important associated plants. Green ephedra, Sandberg bluegrass, Anderson peachbrush, and several forb species also are common. Juniper-pinyon woodland is typical on mountain slopes. Jeffrey pine, lodgepole pine, white fir, and manzanita grow on the highest mountain slopes. Shadscale is the typical plant in the drier parts of the area. Sedges, rushes, and moisture-loving grasses grow on the wettest parts of the wet flood plains and terraces. Basin wildrye, alkali sacaton, saltgrass, buffaloberry, black greasewood, and rubber rabbitbrush grow on the drier sites that have a high concentration of salts.
Some of the major wildlife species in this area are mule deer, coyote, beaver, muskrat, jackrabbit, cottontail, raptors, pheasant, chukar, blue grouse, mountain quail, and mourning dove. The species of fish in the area include trout and catfish. The Lahontan cutthroat trout in the Truckee River is a threatened and endangered species.
LRU notes
The Sierra Influenced Ranges LRU is characterized by wooded great basin mountains with climatic and biotic affinities to the Sierra Nevada mountain range. The Sierra Influences Ranges LRU receives greater precipitation that the mountain ranges of central NV. Amount of precipitation varies in relation to the local strength of the Sierra NV rain shadow, characterized by pinyon and juniper trees. The White, Sweetwater, Pine Nut, Wassuk, and Virginia ranges of Nevada support varying amounts of Sierra Nevada flora, such as ponderosa pine. Elevations range from 1610 to 2420 meters and slopes range from 5 to 49 percent, with a median value of 22 percent. Frost free days (FFD) ranges from 92 to 163.
Ecological site concept
The Mahogany Savanna site occurs on mountain summits and upper side slopes on all exposures. Slopes range from 4 to 50 percent. Elevations are 6000 to 9600 feet. The soils associated with this site are well drained and shallow to very deep. The dominant plants are curl-leaf mountain mahogany (Cercocarpus ledifolius), mountain big sagebrush (Artemisia tridentata ssp. vaseyana), needlegrass (Achnatherum), and bluegrass (Poa).
Associated sites
| R026XY038NV |
LOAMY SLOPE 14+ P.Z. |
|---|---|
| R026XY039NV |
CLAYPAN 14+ P.Z. |
| R026XY081NV |
MAHOGANY THICKET |
Similar sites
| R026XY081NV |
MAHOGANY THICKET Overstory canopy of CELE3 averages more than 50 percent; more productive understory. |
|---|---|
| R026XY080NV |
STONY MAHOGANY SAVANNA Overstory canopy of CELE3 averages more than 50 percent; less productive understory. |
Table 1. Dominant plant species
| Tree |
(1) Cercocarpus ledifolius |
|---|---|
| Shrub |
(1) Artemisia tridentata ssp. vaseyana |
| Herbaceous |
(1) Achnatherum |
Physiographic features
This site occurs on hills, mountain summits and upper side slopes on all exposures. Slopes range from 4 to 50 percent. Elevations are 6000 to over 9600 feet.
Table 2. Representative physiographic features
| Landforms |
(1)
Mountain slope
|
|---|---|
| Elevation | 6,000 – 9,600 ft |
| Slope | 4 – 50% |
| Aspect | Aspect is not a significant factor |
Climatic features
The climate associated with this site is subhumid with cool, dry summers and cold, wet winters. Average annual precipitation is 14 to over 30 inches. Mean annual air temperature is 36 to 42 degrees F. The average growing season is about 50 to 90 days.
Nevada’s climate is predominantly arid, with large daily ranges of temperature, infrequent severe storms, heavy snowfall in the higher mountains, and great location variations with elevation. Three basic geographical factors largely influence Nevada’s climate: continentality, latitude, and elevation. Continentality is the most important factor. The strong continental effect is expressed in the form of both dryness and large temperature variations. Nevada lies on the eastern, lee side of the Sierra Nevada Range, a massive mountain barrier that markedly influences the climate of the State. The prevailing winds are from the west, and as the warm moist air from the Pacific Ocean ascend the western slopes of the Sierra Range, the air cools, condensation occurs and most of the moisture falls as precipitation. As the air descends the eastern slope, it is warmed by compression, and very little precipitation occurs. The effects of this mountain barrier are felt not only in the West but throughout the state, with the result that the lowlands of Nevada are largely desert or steppes. The temperature regime is also affected by the blocking of the inland-moving maritime air. Nevada sheltered from maritime winds, has a continental climate with well-developed seasons and the terrain responds quickly to changes in solar heating.
Nevada lies within the mid-latitude belt of prevailing westerly winds which occur most of the year. These winds bring frequent changes in weather during the late fall, winter and spring months, when most of the precipitation occurs. To the south of the mid-latitude westerlies, lies a zone of high pressure in subtropical latitudes, with a center over the Pacific Ocean. In the summer, this high-pressure belt shifts northward over the latitudes of Nevada, blocking storms from the ocean. The resulting weather is mostly clear and dry during the summer and early fall, with scattered thundershowers. The eastern portion of the state receives significant summer thunderstorms generated from monsoonal moisture pushed up from the Gulf of California, known as the North American monsoon. The monsoon system peaks in August and by October the monsoon high over the Western U.S. begins to weaken and the precipitation retreats southward towards the tropics (NOAA 2004).
Table 3. Representative climatic features
| Frost-free period (characteristic range) | |
|---|---|
| Freeze-free period (characteristic range) | |
| Precipitation total (characteristic range) | 14-30 in |
| Frost-free period (average) | 70 days |
| Freeze-free period (average) | |
| Precipitation total (average) | 22 in |
Figure 1. Monthly precipitation range
Figure 2. Monthly average minimum and maximum temperature
Figure 3. Annual precipitation pattern
Figure 4. Annual average temperature pattern
Influencing water features
There are no influencing water features associated with the Mahogany Savanna site.
Soil features
The soils are well drained and shallow to very deep. Available water capacity is low to moderate because of shallow depth or to high volume of rock fragments in the profile of deeper soils. The soils are slightly acid or neutral. Soil series associated with this site include: Bakscratch, Bradshaw, Fusuvar, Sup and Ticino.
Table 4. Representative soil features
| Parent material |
(1)
Residuum
–
andesite
|
|---|---|
| Surface texture |
(1) Very gravelly loam (2) Extremely stony loam (3) Gravelly fine sandy loam |
| Family particle size |
(1) Loamy |
| Drainage class | Well drained |
| Permeability class | Moderate |
| Depth to restrictive layer | 10 – 59 in |
| Soil depth | 10 – 59 in |
| Surface fragment cover <=3" | 10 – 40% |
| Surface fragment cover >3" | 5 – 39% |
| Available water capacity (0-40in) |
1.1 – 3.4 in |
| Calcium carbonate equivalent (0-40in) |
Not specified |
| Electrical conductivity (0-40in) |
Not specified |
| Sodium adsorption ratio (0-40in) |
Not specified |
| Soil reaction (1:1 water) (0-40in) |
5.6 – 6.5 |
| Subsurface fragment volume <=3" (Depth not specified) |
17 – 31% |
| Subsurface fragment volume >3" (Depth not specified) |
45% |
Ecological dynamics
An ecological site is the product of all the environmental factors responsible for its development and it has a set of key characteristics that influence a site’s resilience to disturbance and resistance to invasive species. Key characteristics include 1) climate (precipitation, temperature), 2) topography (aspect, slope, elevation, and landform), 3) hydrology (infiltration, runoff), 4) soils (depth, texture, structure, organic matter), 5) plant communities (functional groups, productivity), and 6) natural disturbance regime (fire, herbivory, etc.) (Caudle 2013). Biotic factors that influence resilience include site productivity, species composition and structure, and population regulation and regeneration (Chambers et al. 2013).
The Great Basin vegetation communities have high spatial and temporal variability in precipitation both among years and within growing seasons. Nutrient availability is typically low but increases with elevation and closely follows moisture availability. The moisture resource supporting the greatest amount of plant growth is usually the water stored in the soil profile during the winter. The invasibility of plant communities is often linked to resource availability. Disturbance can decrease resource uptake due to damage or mortality of the native species and depressed competition, or increase resource uptake by the decomposition of dead plant material following disturbance. The invasion of cheatgrass (Bromus tectorum) has been linked to disturbances (fire, abusive grazing) that have resulted in fluctuations in resources (Chambers et al. 2007). Dobrowolski et al. (1990) cite multiple authors on the extent of the soil profile exploited by the competitive exotic annual cheatgrass. Specifically, the depth of rooting is dependent on the size the plant achieves; in competitive environments cheatgrass roots were found to penetrate only 15 cm, while isolated plants and pure stands were found to root up to 1.7 m. Mahogany stands are susceptible to drought, frost, and invasion by non-native species, especially cheatgrass. Cheatgrass affects mahogany seedling growth by competing for water resources and nutrients in an area (Ross 1999).
Periodic drought regularly influences Great Basin ecosystems, and drought duration and severity has increased throughout the 20th century in much of the Intermountain West. Major shifts away from historic precipitation patterns have the greatest potential to alter ecosystem function and productivity. Species composition and productivity can be altered by the timing of precipitation and water availability within the soil profile (Bates et al. 2006).
Long-lived curl-leaf mountain mahogany, deep-rooted cool season perennial bunchgrasses, and long-lived shrubs (50+ years) with high root-to-shoot ratios dominate the ecological sites in this DRG. Curl-leaf mountain mahogany (hereafter, mahogany) is a widespread species in Nevada, occurring on mountain slopes spanning a wide elevation range, from the sagebrush and pinyon-juniper vegetation zones into the mountain shrub communities that border mixed conifer and even subalpine ecosystems (Tueller 1989). Mahogany is a multi-branched, evergreen shrub or tree reaching 3 to over 20 feet in height. Mahogany plants are long-lived; it is common to find plants over 200 years old, however there are some reports of plants over 1,000 years old (Ex et al. 2011, Schultz 1987, Schultz et al. 1990). As mahogany stands increase in average age, average canopy volume and height of the individuals present also increases. As average canopy height and volume increase, stand density declines (Schultz et al. 1991). Stands with a closed or nearly closed canopy often have little recruitment in the understory (Schultz et al. 1990, 1991), despite high seed density beneath trees (Russell and Schupp 1998, Ibáñez et al. 2002). Intraspecific competition reduces the growth rates of all age classes and may increase mortality in the younger plants.
The species plays an important role in biogeochemical cycles, since its roots can host nitrogen-fixing nodules (Youngberg and Hu 1972, Freund et al., 2018), possibly allowing for successional processes on poor soils in stressful environments (Kratsch and Graves, 2004). Seedlings of mahogany exhibit rapid root growth in relation to top growth, providing some resistance to drought and competition with invasive species (Dealy 1974). Dealy (1974) reported that curl-leaf mahogany seedlings have a mean taproot length of 0.97 m after 120 days, while the mean top height was slightly less than 2.5 cm. Ibáñez et al. (1999) and Schultz et al. (1996) found that mahogany seedlings germinate abundantly under the canopy of adult plants, but rarely successfully establish there due to shading and higher litter amounts. In addition, Schultz et al. (1996) found that seedlings had significantly higher long term success in areas dominated by sagebrush canopy than in areas under mahogany canopy or in interspaces. The shading and hydraulic lift provided by adult sagebrush may create a microsite that facilitates mahogany recruitment (Gruell et al. 1985, Ibáñez et al. 1999).
Mountain big sagebrush is a minor component of this site. Infrequent large recruitment events and simultaneous low, continuous recruitment is the foundation of population maintenance for this plant (Noy-Meir 1973). It is not tolerant of shade and may be missing in dense stands of mahogany.
Perennial bunchgrasses generally have shallower root systems than shrubs, but root densities are often as high as or higher than those of shrubs in the upper 0.5 m and taper off more rapidly than shrubs. Letterman needlegrass, the dominant grass on the non-modal ecological site, is an erect, densely-tufted perennial bunchgrass that forms large clumps. It is found on dry soils in a variety of vegetation communities, including, high elevation meadows, subalpine grasslands, open areas underneath aspen, and in sagebrush communities. It grows best on loamy soils with greater than 20 cm depth (Dittberner and Olson 1983).
Western needlegrass is a strongly tufted perennial grass that grows up to 4 dm in height (Cronquist et al. 1994). It grows in dry, well-drained soils from upper foothills up into the higher areas of the mountains in the western United States (USDA Forest Service 1988). The roots of this grass are deep, fibrous and spreading, which allows it to be more resistant to trampling and drought (USDA Forest Service 1988).
Cusick’s bluegrass and/or muttongrass are found on this site. There is evidence that these two common names have been used interchangeably or are sometimes misidentified (Monsen et al. 2004), but they occupy similar ecological niches (Cronquist et al. 1972). Cusick’s bluegrass is a strongly tufted perennial grass but may be somewhat rhizomatous in loose soils (Cronquist et al. 1972). It begins growth very early in the season and may produce two crops of inflorescences in a growing season (Cronquist et al. 1972). Muttongrass persists well in open areas and under canopies of oak and other shrubs (Monsen et al. 2004). Muttongrass may be more shade tolerant than other perennial bunchgrasses and may persist in the understory as the canopy closes (Erdman 1970).
There is potential for infilling by singleleaf pinyon (Pinus monophylla) on these sites. Infilling may occur if the site is adjacent to woodland sites or other ecological sites with conifers present. Without disturbance, such as low-intensity fire, pinyon will eventually dominate and out-compete mahogany for water and sunlight. The authors have observed this phenomenon and there is ongoing research to evaluate this process. One study found that mahogany seedlings responded best to mechanical juniper removal compared to burning (O’Connor et al. 2013).
The ecological sites in this DRG have moderate resilience to disturbance and resistance to invasion. Resilience increases with elevation, aspect, precipitation, and nutrient availability. Long-term disturbance response may be influenced by small differences in landscape topography. North slopes are more resilient than south slopes because lower soil surface temperatures operate to keep moisture content higher on northern exposures. Four possible alternative stable states have been identified for this DRG.
Fire Ecology:
The fire return interval in mahogany-dominated sites is not well documented, however a study by Arno and Wilson (1986) suggested mahogany with ponderosa pine communities had fire return intervals of 13 to 22 years before 1900. Fire frequency most likely depends on surrounding vegetation. Most often mahogany stands occur on warm, dry, rocky ridges or outcrops where fire would be an infrequent occurrence (USDA 1937). Dealy (1974) and Scheldt (1969) found that mahogany trees were larger and older on fire-resistant rocky sites and were the seed source if fire destroyed the non-rocky portion of the site. Mahogany is considered a weak sprouter, and is usually moderately to severely damaged by severe fires. Because of their thicker bark, mature trees can often survive low-severity fires (Gruell et al. 1985). The recovery time of these sites is variable; some measurements show that stands lack recruitment for up to 30 years post-fire (Gruell et al. 1985, Ross 1999). Mahogany seeds germinate and have the highest survival rates with moderate litter amounts; litter depths over 0.25 inches can impede recruitment (Gruell et al. 1985, Ibáñez et al. 2002, Ibáñez et al., 1999, Schultz et al. 1991, Schultz et al. 1990). Since these plants germinate well under the protection of adult mahogany and sagebrush, germination rates may be quite low immediately after fire (Shultz et al. 1996, Ross 1999).
The effect of fire on bunchgrasses relates to culm density, culm-leaf morphology, and the size of the plant. The initial condition of bunchgrasses within the site along with seasonality and intensity of the fire all factor into the individual species response. For most forbs and grasses the growing points are located at or below the soil surface providing relative protection from disturbances which decrease above ground biomass, such as grazing or fire. Thus, fire mortality is more correlated to duration and intensity of heat, which is related to culm density, culm-leaf morphology, size of plant and abundance of old growth (Wright 1971, Young 1983). In addition, season and severity of the fire will influence plant response as will post-fire soil moisture availability.
Emergence of western needlegrass seeds was shown to significantly improve with additions of smoke and burned soil (Blank et al. 1996).
Mountain big sagebrush is killed by fire (Neuenschwander 1980, Blaisdell et al. 1982), and does not resprout (Blaisdell 1953). Post fire regeneration occurs from seed and will vary depending on site characteristics, seed source, and fire characteristics. Mountain big sagebrush seedlings can grow rapidly and may reach reproductive maturity within 3 to 5 years (Bunting et al. 1987). Mountain big sagebrush may return to pre-burn density and cover within 15-20 years following fire, but establishment after severe fires may proceed more slowly (Bunting et al. 1987).
Depending on fire severity, snowberry and other sprouting shrubs may increase after fire. Snowberry is top-killed by fire, but resprouts after fire from rhizomes (Leege and Hickey 1971, Noste and Bushey 1987). Snowberry has been noted to regenerate well and exceed pre-burn biomass in the third season after a fire (Merrill et al. 1982). Douglas rabbitbrush (Chrysothamnus viscidiflorus) is also found in these sites. It has a large taproot root system and is known to be shorter lived and less competitive than sagebrush. Seedling density, flower production, and shoot growth decline as competition from other species increases (McKell and Chilcote 1957, Miller et al. 2013). Douglas rabbitbrush is top-killed by fire, but sprouts vigorously after fire (Kuntz 1982, Akinsoji 1988).
Livestock/Wildlife Grazing Interpretations:
Mountain mahogany is an important cover and browse species for big game such as elk (Cervus canadensis), mule deer (Odocoileus heminous), pronghorn antelope (Antilocarpra americana), and bighorn sheep (Ovis canadensis) (Lanner 1984, Furniss et al. 1988, Sabo et al. 2005). Sampson and Jespersen (1963) state that mahogany is excellent browse for mule deer, and domestic livestock will browse this plant to varying degrees in all seasons except summer. It is not uncommon for these trees to develop a “hedged” appearance after years of regular browsing by wildlife. According to (Olsen 1992) mahogany is consumed widely by mule deer throughout the year. In fact, mule deer fecal pellets were observed to contain mahogany year-round, with the highest frequency of leaves found in winter (Gucker 2006). Mule deer will use mahogany for cover as well (Steele et al. 1981).
Despite low palatability, mountain big sagebrush is eaten in small amounts by sheep, cattle, goats, and horses. Chemical analysis indicates that the leaves of big sagebrush equal alfalfa meal in protein, have a higher carbohydrate content, and yield twelvefold more fat (USDA 1937).
Antelope bitterbrush is a small component of these sites, but is a critical browse species for mule deer, antelope and elk and is often utilized heavily by domestic livestock (Wood et al. 1995). Grazing tolerance is dependent on site conditions (Garrison 1953) and the shrub can be severely hedged during the dormant season for grasses and forbs.
Letterman’s needlegrass provides valuable forage for both livestock and wildlife (Taylor 2000). It begins growth early in the year and is available to be utilized when other grasses are not yet palatable. It is especially important fall forage for big game (Monsen et al. 2004). Letterman’s needlegrass appears to tolerate sheep grazing, however time and timing of grazing is not well documented for this species (Bowns and Bagley 1986). It also declines when grazing is excluded for a long time (Turner 1969).
Western needlegrass is slow to mature and remains green through most of the growing season. Since it can remain green into fall, it is higher quality forage compared to other species that have senesced by then (USDA Forest Service 1988). For livestock this grass has good forage value, and it has fair forage value for wildlife (Stubbendieck et al. 1992). Seeds of this grass are avoided by grazing animals, but are not necessarily injurious. Since seeds are avoided by grazing animals, a large amount of the seed produced grows to maturity (USDA Forest Service 1988).
Cusick’s bluegrass was the most palatable and preferred grass compared to Thurber’s needlegrass and bluebunch wheatgrass in a 1975 grazing study, and was the most negatively affected by grazing (Rickard et al. 1975). Uresk and Rickard (1976) found Cusick’s bluegrass to be a highly preferred grass, especially in the spring, even when it is a minor component of the plant community.
Invasive Annual Grasses:
The species most likely to invade these sites is cheatgrass. Cheatgrass is a cool season annual grass that maintains an advantage over native plants for several reasons: it is a prolific seed producer, can germinate in the autumn or spring, tolerates grazing, and increases with frequent fire (Klemmedson and Smith 1964, Miller et al. 1999). Cheatgrass originated from Eurasia and was first reported in North America in the late 1800s (Mack and Pyke 1983; Furbush 1953). Pellant and Hall (1994) found 3.3 million acres of public lands dominated by cheatgrass and suggested that another 76 million acres were susceptible to invasion by winter annuals including cheatgrass and medusahead.
Recent modeling and empirical work by Bradford and Lauenroth (2006) suggests that seasonal patterns of precipitation input and temperature are also key factors determining regional variation in the growth, seed production, and spread of invasive annual grasses. The phenomenon of cheatgrass “die-off” provides opportunities for restoration of perennial and native species (Baughman et al. 2016, Baughman et al. 2017). The causes of these events are not fully understood, but there is ongoing work to try to predict where they occur, in the hopes of aiding conservation planning (Weisberg et al. 2017, Brehm 2019).
Methods to control cheatgrass include herbicide, fire, targeted grazing, and seeding. Mapping potential or current invasion vectors is a management method designed to increase the cost effectiveness of control methods. Spraying with herbicide (Imazapic or Imazapic + glyphosate) and seeding with crested wheatgrass and Sandberg bluegrass has been found to be more successful at combating cheatgrass (and medusahead) than spraying alone (Sheley et al. 2012). To date, most seeding success has occurred with non-native wheatgrass species. Perennial grasses, especially crested wheatgrass, are able to suppress cheatgrass growth when mature (Blank et al. 2020). Where native bunchgrasses are missing from the site, revegetation of annual grass invaded rangelands has been shown to have a higher likelihood of success when using introduced perennial bunchgrasses such as crested wheatgrass (Clements et al. 2017, Davies et al. 2015). Butler et al. (2011) tested four herbicides (Imazapic, Imazapic + glyphosate, rimsulfuron, and sulfometuron + Chlorsulfuron) for suppression of cheatgrass, medusahead and ventenata (North Africa grass, Ventenata dubia) within residual stands of native bunchgrass. Additionally, they tested the same four herbicides followed by seeding of six bunchgrasses (native and non-native) with varying success (Butler et al. 2011). Herbicide-only treatments appeared to remove competition for established bluebunch wheatgrass by providing 100% control of ventenata and medusahead and greater than 95% control of cheatgrass (Butler et al. 2011). Caution in using these results is advised, as only one year of data was reported.
In considering the combination of pre-emergent herbicide and prescribed fire for invasive annual grass control, it is important to assess the tolerance of desirable brush species to the herbicide being applied. Vollmer and Vollmer (2008) tested the tolerance of alderleaf mountain mahogany (Cercocarpus montanus), antelope bitterbrush, and multiple sagebrush species to three rates of Imazapic with and without methylated seed oil as a surfactant. They found a cheatgrass control program in an antelope bitterbrush community should not exceed Imazapic at 8 oz./ac with or without surfactant. Sagebrush, regardless of species or rate of application, was not affected. However, many environmental variables were not reported in this study and managers should install test plots before broad scale herbicide application is initiated.
State and Transition Model Narrative for Group 15:
This is a text description of the states, phases, transitions, and community pathways possible in the State and Transition model for the MLRA 26 Disturbance Response Group 15. Site included in this Disturbance Response Group are R026XY009NV and R026XY015CA.
Reference State 1.0:
The Reference State 1.0 represents the natural range of variability under pristine conditions. The reference state has three general community phases; a shrub-grass dominant phase, a perennial grass dominant phase and a shrub dominant phase. State dynamics are maintained by interactions between climatic patterns and disturbance regimes. Negative feedbacks enhance ecosystem resilience and contribute to the stability of the state. These include the presence of all structural and functional groups, low fine fuel loads, and retention of organic matter and nutrients. Plant community phase changes are primarily driven by fire, periodic drought and/or insect or disease attack.
Community Phase 1.1:
This community is dominated by curl-leaf mountain mahogany. Mountain big sagebrush and snowberry make up the shrub components of the understory. Needlegrasses and bluegrasses are dominant perennial bunchgrasses. A diversity of other grasses and forbs exist in the understory.
Community Phase Pathway 1.1a from phase 1.1 to 1.2:
Fire will reduce the mahogany overstory and allow the understory species to dominate the site. Due to low fuel loads, fires will typically be low severity, resulting in a mosaic pattern.
Community Phase Pathway 1.1b from phase 1.1 to 1.3:
Time and lack of disturbance or fire, drought, herbivory, or combinations of these causes mountain mahogany to increase. The shrub and herbaceous understory components decline due to increased shading from the trees. Muttongrass increases with more shade.
Community Phase 1.2:
This community phase is characteristic of a post-disturbance, early- to mid-seral plant community. Snowberry and rabbitbrush are sprouting. Perennial grasses and forbs dominate. Mahogany and mountain big sagebrush may be present, but only in patches.
Community Phase Pathway 1.2a from phase 1.2 to 1.1:
Time and lack of disturbance or fire, drought, herbivory, or combinations of these allows the mountain mahogany and sagebrush to increase.
Community Phase 1.3 (At-Risk):
Mahogany density will increase in the absence of disturbance. Shrubs and deep-rooted perennial bunchgrasses will be shaded out by the dense mahogany. Bluegrasses are more shade tolerant, however, and increase in the understory. Mahogany in dense stands will lose lower branches due to shading and/or herbivory, resulting in a more tree-like appearance.
Community Phase Pathway 1.3a fro, phase 1.3 to 1.2:
A low-severity or spot fire, snow loading, or insect damage will decrease the overstory and allow for the herbaceous plants in the understory to increase.
T1A: Transition from Reference State 1.0 to Current Potential State 2.0:
Trigger: This transition is caused by the introduction of non-native annual weeds, such as cheatgrass, mustard and Russian thistle.
Slow variables: Over time, the annual non-native plants will increase within the community decreasing organic matter inputs from deep-rooted perennial bunchgrasses resulting in reductions in soil water availability for perennial bunchgrasses.
Threshold: Any amount of introduced non-native species causes an immediate decrease in the resilience of the site. Annual non-native species cannot be easily removed from the system and have the potential to significantly alter disturbance regimes from their historic range of variation.
T1B: Transition from Reference State 1.0 to Tree State 3.0:
Trigger: Time and lack of disturbance allows pinyon to increase and overtop the mountain mahogany. Litter increases while understory plants decrease.
Slow variables: Over time, abundance and size of singleleaf pinyon will increase.
Threshold: Pinyon dominate(s) ecological processes. Trees overtop and outcompete mountain mahogany and shrubs for water and sunlight. Shrub skeletons exceed live shrubs with minimal recruitment of new cohorts.
Current Potential State 2.0:
This state is similar to the Reference State 1.0. Ecological function has not changed, however the resiliency of the state has been reduced by the presence of invasive weeds. This state has the same three general community phases. Negative feedbacks enhance ecosystem resilience and contribute to the stability of the state. These include the presence of all structural and functional groups, low fine fuel loads and retention of organic matter and nutrients. Positive feedbacks decrease ecosystem resilience and stability of the state. These include the non-natives high seed output, persistent seed bank, rapid growth rate, ability to cross-pollinate and adaptations for seed dispersal. Additionally, the presence of highly flammable, non-native species reduces State resilience because these species can promote fire where historically fire has been infrequent leading to positive feedbacks that further the degradation of the system.
Community Phase 2.1:
This community is dominated by curl-leaf mountain mahogany. Mountain big sagebrush and snowberry make up the shrub components of the understory. Needlegrasses and bluegrasses are dominant perennial bunchgrasses. A diversity of other grasses and forbs exist in the understory. Annual non-native species like cheatgrass are present.
Community Phase Pathway 2.1a from phase 2.1 to 2.2:
Fire will decrease or eliminate the overstory of mahogany and allow the perennial bunchgrasses to dominate the site. Fires will typically be small and patchy due to low fuel loads.
Community Phase Pathway 2.1b from phase 2.1 to 2.3:
Time and lack of disturbance or fire, drought, herbivory, or combinations of these causes mountain mahogany to increase. The shrub and herbaceous understory components decline due to increased shading from the mahogany and/or pinyon pine. Muttongrass increases with more shade.
Community Phase 2.2:
This community phase is characteristic of a post-disturbance, early to mid-seral community phase. Needlegrasses and other perennial grasses dominate the site. Snowberry and/or rubber rabbitbrush may be sprouting. Mountain mahogany and mountain big sagebrush are patchy. Annual non-native species are present.
Community Phase Pathway 2.2a from phase 2.2 to 2.1:
Time and lack of disturbance or fire, drought, herbivory, or combinations of these allows the mountain mahogany and sagebrush to increase.
Community Phase Pathway 2.2b from phase 2.2 to 2.4:
Higher than normal spring precipitation favors annual non-native species such as cheatgrass. Non-native annual species will increase in production and density throughout the site. Perennial bunchgrasses may also increase in production. Fire may also play a part in this pathway.
Community Phase 2.3 (At-Risk):
Mahogany density will increase in the absence of disturbance. Shrubs and deep-rooted perennial bunchgrasses will be shaded out by the dense mahogany. Bluegrasses are more shade tolerant, however, and increase in the understory. Mahogany in dense stands will lose lower branches due to shading and/or herbivory, resulting in a more tree-like appearance. Pinyon pine may be present.
Community Phase Pathway 2.3a from phase 2.3 to 2.2:
Fire reduces the shrub overstory and allows perennial bunchgrasses to dominate the site. Fires are typically low severity resulting in a mosaic pattern due to low fuel loads. A fire following an unusually wet spring or a change in management favoring an increase in fine fuels may be more severe and reduce sagebrush cover to trace amounts. Annual non-native species are likely to increase after fire.
Community Phase Pathway 2.3b from phase 2.3 to 2.4:
Fall, winter, and spring precipitation and temperatures mediate the ability for annual grasses and perennial grasses to germinate and/or survive. Higher than normal spring precipitation creates high annual production of annual grasses (Bradley et al. 2016). Non-native annual species increase in production and density throughout the site. Perennial bunchgrasses may also increase in production.
Community Phase 2.4 (At-Risk):
This community is at risk of crossing into an annual state. Native bunchgrasses dominate; however, annual non-native species such as cheatgrass may be sub- or co-dominant in the understory. Annual production and abundance of these annuals may increase drastically in years with heavy spring precipitation. This site is susceptible to further degradation from grazing, drought, and fire. Pinyon pine may be present.
T2A: Transition from Current Potential State 2.0 to Tree State 3.0:
Trigger: Time and lack of disturbance allows pinyon to increase and overtop the mountain mahogany. Litter increases while understory plants decrease.
Slow variables: Over time, abundance and size of pinyon will increase.
Threshold: Pinyon pine dominate(s) ecological processes. Trees overtop and outcompete mountain mahogany and shrubs for water and sunlight. Shrub skeletons exceed live shrubs with minimal recruitment of new cohorts.
T2B: Transition from Current Potential State 2.0 to Annual State 4.0:
Trigger: Fire or a failed range seeding leads to plant community phase 4.1. Inappropriate grazing management that favors shrubs in the presence of non-native annual species leads to community phase 4.2.
Slow variables: Increased production and cover of non-native annual species.
Threshold: Cheatgrass or other non-native annuals dominate understory.
Tree State 3.0:
This state has two community phases that are characterized by the dominance of singleleaf pinyon in the overstory. Mountain big sagebrush and perennial bunchgrasses may still be present, but they are no longer controlling site resources. Soil moisture, soil nutrients, soil organic matter distribution and nutrient cycling have been spatially and temporally altered.
Community Phase 3.1:
Pinyon pine and mountain mahogany dominate the site. Mountain big sagebrush and snowberry are minor component. Bluegrasses dominate understory. Annual non-native species may be present or dominant.
Community Phase Pathway 3.1a from phase 3.1 to 3.2:
Time and lack of disturbance or fire, drought, inappropriate grazing management, or combinations of these allows for maturation of the pinyon/juniper community.
Community Phase 3.2:
Pinyon pine dominates the site. Mountain mahogany is decadent and the stand lacks recruitment. Bluegrasses are present. Understory is reduced overall. Annual non-native species may be present.
T3A: Transition from Tree State 3.0 to Annual State 4.0:
Trigger: To community phase 4.1: Overgrazing in the presence of non-native annual species can cause a decrease in perennial bunchgrasses and an increase in annual species. Spring and/or fall moisture may also increase annual species. To community phase 4.2: Fire in the presence of annual invasive grasses.
Slow variables: Cover and production of annual non-native species increase in the understory.
Threshold: Loss of mahogany overstory, mountain big sagebrush, and deep-rooted perennial bunchgrasses changes nutrient cycling, nutrient redistribution, and reduces soil organic matter. Increased, continuous fine fuels modify the fire regime by changing intensity, size and spatial variability of fires.
R3A: Restoration from Tree State 3.0 to Current Potential State 2.0:
Removal of pinyon from site will allow mountain mahogany to again become the dominant overstory.
Annual State 4.0:
This state has two community phases: one with annual invasive plants in the understory of an intact mahogany stand, and another post-fire phase where mahogany is a minor component or missing from the site. This state is characterized by the dominance of annual non-native species such as cheatgrass and/or tansy mustard in the understory. Ecological dynamics are significantly altered in this state. Annual non-native species create a highly combustible fuel bed that shortens the fire return interval. Nutrient cycling is spatially and temporally truncated as annual plants contribute significantly less to deep soil carbon. Because this is a productive site, some deep-rooted perennial grasses may remain, even in the annual state. Without management, it is unlikely these plants will be able to recruit in the presence of dominant annual grasses.
Community Phase 4.1:
Mountain mahogany dominates the overstory and annual non-native plants such as cheatgrass dominate the understory. Native perennial grasses and forbs are significantly reduced. Sagebrush and snowberry may or may not be present.
Community Phase Pathway 4.1a:
Catastrophic fire reduces the mountain mahogany overstory and allows annual species to dominate.
Community Phase 4.2:
Annual non-native species dominate the site. The open canopy may allow sprouting shrubs and bluegrasses to increase.
State and transition model
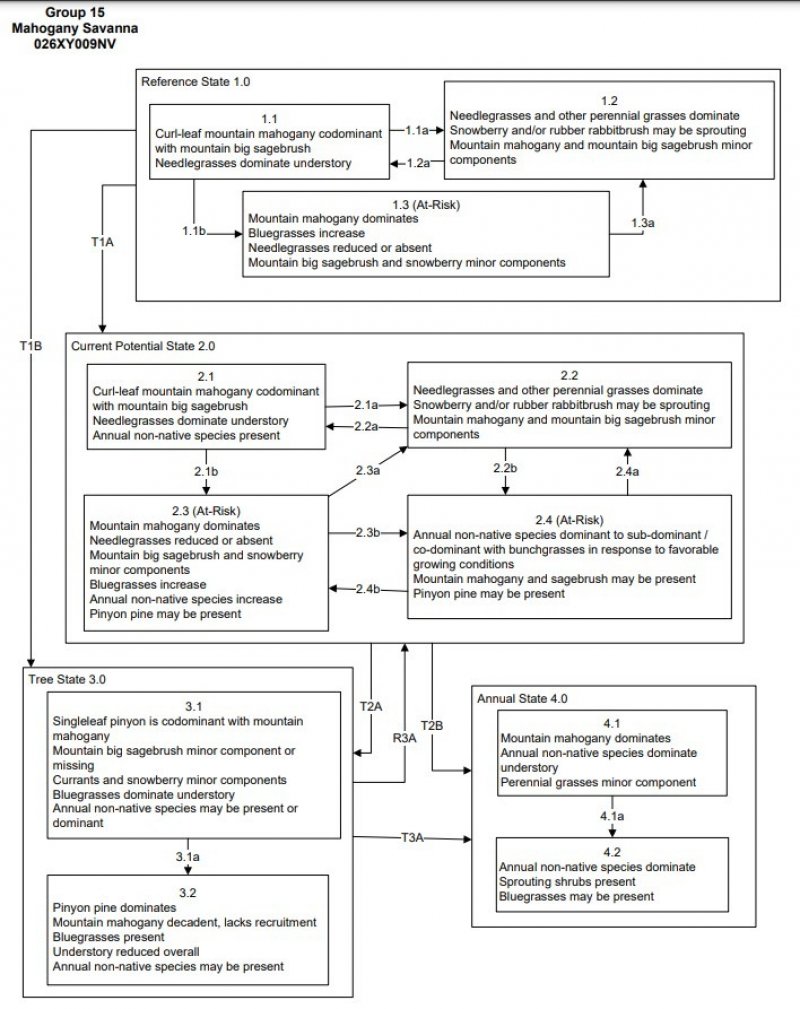

More interactive model formats are also available.
View Interactive Models
Click on state and transition labels to scroll to the respective text
Ecosystem states
State 1 submodel, plant communities
State 1
Reference Plant Community
Community 1.1
Reference Plant Community
The reference plant community is dominated by curlleaf mountainmahogany. Mountain big sagebrush, spike fescue, and Letterman's and western needlegrasses are other important species associated with this site. Potential vegetative composition is about 20% grasses, 5% forbs and 75% shrubs and tree-like shrubs. Approximate crown cover of woody plants ranges from 25 percent to about 45 percent. Total annual air-dry production for all vegetation in the understory to a height of 4.5 feet is 700 lbs for favorable years, 900 lbs for normal years and 1300 lbs for favorable years.
Figure 5. Annual production by plant type (representative values) or group (midpoint values)
Table 5. Annual production by plant type
| Plant type | Low (lb/acre) |
Representative value (lb/acre) |
High (lb/acre) |
|---|---|---|---|
| Tree | 2275 | 2925 | 4225 |
| Grass/Grasslike | 700 | 900 | 1300 |
| Shrub/Vine | 350 | 450 | 650 |
| Forb | 175 | 225 | 325 |
| Total | 3500 | 4500 | 6500 |
Additional community tables
Table 6. Community 1.1 plant community composition
| Group | Common name | Symbol | Scientific name | Annual production (lb/acre) | Foliar cover (%) | |
|---|---|---|---|---|---|---|
|
Grass/Grasslike
|
||||||
| 1 | Primary Perennial Grasses | 226–630 | ||||
| Letterman's needlegrass | ACLE9 | Achnatherum lettermanii | 68–180 | – | ||
| western needlegrass | ACOCO | Achnatherum occidentale ssp. occidentale | 68–180 | – | ||
| spike fescue | LEKI2 | Leucopoa kingii | 1–90 | – | ||
| Cusick's bluegrass | POCU3 | Poa cusickii | 30–60 | – | ||
| muttongrass | POFE | Poa fendleriana | 30–60 | – | ||
| 2 | Secondary Perennial Grasses | 1–90 | ||||
| pine needlegrass | ACPI2 | Achnatherum pinetorum | 23–90 | – | ||
| mountain brome | BRMA4 | Bromus marginatus | 23–90 | – | ||
| prairie Junegrass | KOMA | Koeleria macrantha | 23–90 | – | ||
| basin wildrye | LECI4 | Leymus cinereus | 23–90 | – | ||
| rock melicgrass | MEST | Melica stricta | 23–90 | – | ||
| Sandberg bluegrass | POSE | Poa secunda | 23–90 | – | ||
|
Forb
|
||||||
| 3 | Perennial | 90–180 | ||||
| tapertip hawksbeard | CRAC2 | Crepis acuminata | 23–90 | – | ||
| lupine | LUPIN | Lupinus | 23–90 | – | ||
|
Shrub/Vine
|
||||||
| 4 | Primary Shrubs | 90–360 | ||||
| mountain big sagebrush | ARTRV | Artemisia tridentata ssp. vaseyana | 90–360 | – | ||
| 5 | Secondary Shrubs | 90–360 | ||||
| desert sweet | CHMI2 | Chamaebatiaria millefolium | 45–90 | – | ||
| yellow rabbitbrush | CHVI8 | Chrysothamnus viscidiflorus | 45–90 | – | ||
| mormon tea | EPVI | Ephedra viridis | 45–90 | – | ||
| rockspirea | HODU | Holodiscus dumosus | 45–90 | – | ||
| antelope bitterbrush | PUTR2 | Purshia tridentata | 45–90 | – | ||
| currant | RIBES | Ribes | 45–90 | – | ||
| mountain snowberry | SYOR2 | Symphoricarpos oreophilus | 45–90 | – | ||
|
Tree
|
||||||
| 6 | Evergreen | 3150–3600 | ||||
| curl-leaf mountain mahogany | CELE3 | Cercocarpus ledifolius | 3150–3600 | – | ||
Interpretations
Animal community
Livestock Interpretations:
This is generally not an important site for livestock grazing, because of thick stands of tree-form curlleaf mountainmahogany, steep slopes and rockiness of the slopes. This site may be used by livestock in conjunction with associated sites. Muttongrass is excellent forage for domestic livestock especially in the early spring. Muttongrass begins growth in late winter and early spring, which makes it available before many other forage plants. Cusick's bluegrass makes up only a small proportion of the biomass of the sagebrush communities in which it lives, but it is often taken preferentially by cattle, especially early in the season. Domestic livestock commonly graze spike fescue on spring range. Spike fescue is a highly nutritious, productive, and palatable grass. It is fairly palatable for cattle and domestic sheep in the spring; however, as spike fescue matures in summer it becomes unpalatable and is grazed sparingly. Some livestock (domestic goats, sheep, and cattle) use it in spring, fall, and/or winter but rarely in the summer.
Stocking rates vary over time depending upon season of use, climate variations, site, and previous and current management goals. A safe starting stocking rate is an estimated stocking rate that is fine tuned by the client by adaptive management through the year and from year to year.
Wildlife Interpretations:
Deer readily utilize curlleaf mountainmahogany during the winter and spring. Curlleaf mountainmahogany stands provide hiding and thermal cover for a variety of wildlife species. Non-game and game species of birds and small mammals also use this site for cover. Distance from water limits the extent to which this site is used. Mountain big sagebrush is highly preferred and nutritious winter forage for mule deer and elk. Spike fescue is frequently browsed by mule deer and elk. Spike fescue provides some cover for smaller mammals and birds. Deer and elk make heavy use of muttongrass, especially in early spring when other green forage is scarce. Depending upon availability of other nutritious forage, deer may use mutton grass in all seasons. Mutton grass cures well and is an important fall and winter deer food in some areas. Deer, elk, and mountain goat also use Cusick's bluegrass early in the season. The value of Cusick's bluegrass as cover for small animals has been rated as poor to fair. Needlegrass is another important forage species for several wildlife species.
Hydrological functions
Runoff is medium to very high.
Recreational uses
This site has limited value for recreational uses, due to steep slopes and dense canopy of curlleaf mountainmahogany.
Wood products
This site has some value for producing firewood. Curlleaf mountainmahogany is extremely hard to cut. Curlleaf mountainmahogany is renown for its qualities for smoking and barbecuing meats.
Other products
Native Americans used big sagebrush leaves and branches for medicinal teas, and the leaves as a fumigant. Bark was woven into mats, bags and clothing.
Other information
Curlleaf mountainmahogany may be planted to help stabilize soil in disturbed areas such as roadcuts and mine spoils.
Supporting information
Inventory data references
NV-ECS-1
Type locality
| Location 1: Carson City County, NV | |
|---|---|
| General legal description | This site also occurs in Douglas, Lyon, Mineral, Storey and Washoe counties, Nevada. |
Other references
Abbott, M. L., et al. 1991. Root profiles of selected cold desert shrubs and grasses in disturbed and undisturbed soils. Environmental and Experimental Botany 31(2): 165-178.
Akinsoji, A. 1988. Postfire vegetation dynamics in a sagebrush steppe in southeastern Idaho, USA. Vegetatio 78(3):151-155.
Arno, S. F., Wilson, A. E. 1986. Dating past fires in curl-leaf mountain-mahogany communities. Journal of Range Management 39(3): 241-243.
Bates, J. D., R. F. Miller, and K. W. Davies. 2006. Restoration of quaking aspen woodlands invaded by western juniper. Rangeland Ecology and Management 59:88-97.
Baughman, O. W., R. Burton, M. Williams, P. J. Weisberg, T. E. Dilts, and E. A. Leger. 2017. Cheatgrass die-offs: a unique restoration opportunity in northern Nevada. Rangelands 39(6):165-173.
Baughman, O. W., S. E. Meyer, Z. T. Aanderud, and E. A. Leger. 2016. Cheatgrass die-offs as an opportunity for restoration in the Great Basin, USA: Will local or commercial native plants succeed where exotic invaders fail? Journal of Arid Environments 124:193-204.
Bentz, B., D. Alston, and T. Evans. 2008. Great Basin insect outbreaks. Pages 45-48 in Collaborative Management and Research in the Great Basin -- Examining the issues and developing a framework for action U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fort Collins, CO.
Blaisdell, J. P. 1953. Ecological effects of planned burning of sagebrush-grass range on the upper Snake River Plains. US Dept. of Agriculture.
Blaisdell, J. P., R. B. Murray, and E. D. McArthur. 1982. Managing intermountain rangelands -- sagebrush-grass ranges. U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station, Ogden, UT.
Blank, R. R., C. Clements, T. Morgan, D. Harmon, and F. Allen. 2020. Suppression of cheatgrass by perennial bunchgrasses. Rangeland Ecology & Management 73(6):766-771.
Blank, R. R., F. L. Allan, and J. A. Young. 1996. Influence of simulated burning of soil-litter from low sagebrush, squirreltail, cheatgrass, and medusahead on water-soluble anions and cations. International Journal of Wildland Fire 6:137-143.
Bowns, J. E., and C. F. Bagley. 1986. Vegetation responses to long term sheep grazing on mountain ranges. Journal of Range Management 39.
Bradford, J. B. and W. K. Lauenroth. 2006. Controls over invasion of Bromus tectorum: The importance of climate, soil, disturbance and seed availability. Journal of Vegetation Science 17(6): 693-704.
Brehm, J. R. 2019. Cheatgrass die-off in the Great Basin: A comparison of remote sensing detection methods and identification of environments favorable to die-off. M.S. Thesis. University of Nevada, Reno.
Bunting, S. C., B. M. Kilgore, and C. L. Bushey. 1987. Guidelines for prescribed burning sagebrush-grass rangelands in the northern Great Basin. US Department of Agriculture, Forest Service, Intermountain Research Station Ogden, UT, USA.
Butler, M., F. Brummer, J. Weber, and R. Simmons. 2011. Restoring Central Oregon Rangeland from Ventenata and Medusahead to a Sustainable Bunchgrass Environment – Warm Springs and Ashwood. Central Oregon Agriculture Research and Extension Center.
Caudle, D., J. DiBenedetto, M. Karl, H. Sanchez, and C. Talbot. 2013. Interagency ecological site handbook for rangelands.
Chambers, J. C., B. A. Bradley, C. S. Brown, C. D’Antonio, M. J. Germino, J. B. Grace, S. P. Hardegree, R. F. Miller, and D. A. Pyke. 2013. Resilience to stress and disturbance, and resistance to Bromus tectorum L. invasion in cold desert shrublands of western North America. Ecosystems 17:360-375.
Chambers, J. C., B. A. Roundy, R. R. Blank, S. E. Meyer, and A. Whittaker. 2007. What makes great basin sagebrush ecosystems invasible by Bromus tectorum? Ecological Monographs 77:117-145.
Clements, C. D., D. N. Harmon, R. R. Blank, and M. Weltz. 2017. Improving seeding success on cheatgrass-infested rangelands in northern Nevada. Rangelands 39(6):174-181.
Comstock, J. P. and J. R. Ehleringer. 1992. Plant adaptation in the Great Basin and Colorado Plateau. The Great Basin Naturalist 52:195-215.
Cronquist, A., A. H. Holmgren, N. H. Holmgren, and J. H. Reveal. 1972. Intermountain Flora: Vascular Plants of the Intermountain West, U.S.A. Hafner Publishing Company, Inc. , New York and London.
Cronquist, A., A. H. Holmgren, N. H. Holmgren, J. L. Reveal, and P. K. Holmgren. 1994. Intermountain Flora Vascular Plants of the Intermountain West, U.S.A. The New York Botanical Garden, Bronx, New York.
Davies, K. W., C. S. Boyd, D. D. Johnson, A. M. Nafus, and M. D. Madsen. 2015. Success of seeding native compared with introduced perennial vegetation for revegetating medusahead-invaded sagebrush rangeland. Rangeland Ecology & Management 68(3):224-230.
Dealy, J. E. 1974. Ecology of curlleaf mountain-mahogany (Cercocarpus ledifolius Nutt.) in eastern Oregon and adjacent areas. Unpublished dissertation. Corvallis, OR, Oregon State University.
Dittberner, P. L., and M. R. Olson. 1983. The plant information network (PIN) data base: Colorado, Montana, North Dakota, Utah, and Wyoming. Aspen Bibliography.
Dobrowolski, J. P., M. M. Caldwell, and J. H. Richards. 1990. Basin hydrology and plant root systems. Pages 243-292 in C. B. Osmond, L. F. Pitelka, and G. M. Hidy, editors. Plant biology of the basin and range. Springer-Verlag, New York.
Erdman, J. A. 1970. Pinyon-juniper succession after natural fires on residual soils of Mesa Verde, Colorado. Brigham Young University Science Bulletin-Biological Series 11:1-26.
Ex, S. A., R. J. DeRose, and J. N. Long. 2011. Stand development and population dynamics of curlleaf mountain mahogany (Cercocarpus ledifolius Nutt.) woodlands in Utah's Bear River Mountains. Western Journal of Applied Forestry 26:183-188.
Freund, S. M., F. M. Soper, S. R. Poulson, P. C. Selmants, and B. W. Sullivan. 2018. Actinorhizal species influence plant and soil nitrogen status of semiarid shrub-dominated ecosystems in the western Great Basin, USA. Journal of Arid Environments 157:48-56.
Furbush, P. 1953. Control of Medusa-Head on California Ranges. Journal of Forestry 51(2): 118-121.
Furniss, M. M. and W. F. Barr. 1975. Insects affecting important native shrubs of the northwestern United States. US Intermountain Forest And Range Experiment Station. USDA Forest Service General Technical Report INT INT-19.
Furniss, M. M., D. C Ferguson, K. W Voget, J. W. Burkhardt, A.R. Tiedemann, J. L. Oldemeyer (1988). Taxonomy, life history, and ecology of a mountain-mahogany defoliator, Stamnodes animata (Pearsall), in Nevada. Fish and Wildlife Research 3. Washington, DC, U.S. Department of the Interior, Fish and Wildlife Service: 26.
Ganskopp, D., L. Aguilera, and M. Vavra. 2007. Livestock forage conditioning among six northern Great Basin grasses. Rangeland Ecology & Management 60:71-78.
Garrison, G. A. 1953. Effects of clipping on some range shrubs. Journal of Range Management 6:309-317.
Goodrich, S., E. D. McArthur, and A. H. Winward. 1985. A new combination and a new variety in Atemisia tridentata. The Great Basin Naturalist 45:99-104.
Gruell, G., Bunting, S. C., Neuenschwander, L. 1985. Influence of fire on curlleaf mountain-mahogany in the Intermountain West. Fire's Effects on Wildlife Habitat - symposium proceedings. General Technical Report INT-186, Missoula, Montana, USDA Forest Service, Intermountain Research Station.
Gucker, C. L. (2006). Cercocarpus ledifolius. Fire Effects Information System, [Online]. , U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory
Harris, G. A. 1967. Some Competitive Relationships between Agropyron spicatum and Bromus tectorum. Ecological Monographs 37(2): 89-111.
Hironaka, M. 1994. Medusahead: natural successor to the cheatgrass type in the northern Great Basin. Pages 89-91 in Proceedings of Ecology and Management of Annual Rangelands. Gen. Tech. Report INT-313. USDA Forest Service, Intermountain Research Station., Boise, ID.
Humphrey, L. D. 1984. Patterns and mechanisms of plant succession after fire on Artemisia-grass sites in southeastern Idaho. Vegetation 57:91-101.
Ibáñez, I., Eugene, W., Schupp. 2002. Effects of litter, soil surface conditions, and microhabitat on Cercocarpus ledifolius Nutt. seedling emergence and establishment. Journal of Arid Environments 52(2): 209-222.
Ibáñez, I., Schupp, E. W., Boettinger, J. L. (1999). Successional history of a curlleaf mountain mahogany stand: a hypothesis. Proceedings: shrubland ecotones, August 12-14, 1998, Ephraim, UT. E. D. McArthur, W. K.Ostler and C. L. Wambolt, U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Ogden, UT. P-RMRS-P-11. P-RMRS-P-11: 12-14.
Johnson, J. R. and G. F. Payne. 1968. Sagebrush reinvasion as affected by some environmental influences. Journal of Range Management 21:209-213.
Klemmedson, J. O. and J. G. Smith. 1964. Cheatgrass (Bromus Tectorum L.). The botanical review 30(2): 226-262.
Krall, J. L., J. R. Stroh, C. S. Cooper, and S. R. Chapman. 1971. Effect of time and extent of harvesting basin wildrye. Journal of Range Management:414-418.
Kratsch, H. A., and W. R. Graves. 2004. Nitrogen fixation as a stress-avoidance strategy among actinorhizal (non-legume) trees and shrubs. Journal of Crop Improvement 10(1-2):281-304.
Kuntz, D. E. 1982. Plant response following spring burning in an Artemisia tridentata subsp. vaseyana/Festuca idahoensis habitat type. Dissertation, University of Idaho, Moscow, ID.
Lanner, R. M. 1984. Trees of the Great Basin: A Natural History, University of Nevada Press.
Leege, T. A., and W. O. Hickey. 1971. Sprouting of northern Idaho shrubs after prescribed burning. The Journal of Wildlife Management 35(3):508-515.
Lett, M. S. and A. K. Knapp. 2005. Woody plant encroachment and removal in mesic grassland: production and composition responses of herbaceous vegetation. The American Midland Naturalist 153:217-231.
Mack, R. N. and D. Pyke. 1983. The demography of Bromus Tectorum: Variation in time and space. Journal of Ecology 71(1): 69-93.
Majerus, M. E. 1992. High stature grasses for winter grazing. Journal of Soil and Water Conservation 47:224-225.
Mangla, S., R. Sheley, and J. J. James. 2011. Field growth comparisons of invasive alien annual and native perennial grasses in monocultures. Journal of Arid Environments 75(2):206-210.Monsen, S. B. 1992. The competitive influences of cheatgrass (Bromus tectorum) on site restoration. Pages 43-50 in Proceedings - Ecology, Management, and Restoration of Intermountain Annual Rangelands. General Technical Report INT-GTR-313. U.S.D.A Forest Service Intermountain Research Station, Boise, ID.
McKell, C. M., and W. W. Chilcote. 1957. Response of rabbitbrush following removal of competing vegetation. Journal of Range Management Archives 10(5):228-229.
Merrill, E. H., H. Mayland, and J. Peek. 1982. Shrub responses after fire in an idaho ponderosa pine community. The Journal of Wildlife Management 46(2):496-502.
Miller, H. C., D. Clausnitzer, and M. M. Borman. 1999. Medusahead. Pages 272-281 in R. L. Sheley and J. K. Petroff, editors. Biology and Management of Noxious Rangeland Weeds. Oregon State University Press, Corvallis, OR.
Miller, R. F. and E. K. Heyerdahl. 2008. Fine-scale variation of historical fire regimes in sagebrush-steppe and juniper woodland: an example from California, USA. International Journal of Wildland Fire 17:245-254
Miller, R. F. and R. J. Tausch. 2000. The role of fire in pinyon and juniper woodlands: a descriptive analysis. Pages 15-30 in Proceedings of the invasive species workshop: the role of fire in the control and spread of invasive species. Fire conference.
Miller, R. F., J. C. Chambers, D. A. Pyke, F. B. Pierson, and C. J. Williams. 2013. A Review of Fire Effects on Vegetation and Soils in the Great Basin Region: Response and Ecological Site Characteristics. USDA Forest Service, Rocky Mountain Research Station Fort Collins, CO. 126 p.
Monsen, S. B., R. Stevens, and N. L. Shaw. 2004. Grasses. Pages 295-424 in Restoring western ranges and wildlands, vol. 2. Gen. Tech. Rep. RMRS-GTR-136-vol-2. USDA: Forest Service, Rocky Mountain Research Station, Fort Collins, CO.
Neuenschwander, L. F. 1980. Broadcast burning of sagebrush in the winter. Journal of Range Management 33:233-236.
Neuenschwander, L. F. 1980. Broadcast burning of sagebrush in the winter. Journal of Range Management 33(3):233-236.
Noste, N. V., and C. L. Bushey. 1987. Fire response of shrubs of dry forest habitat types in Montana and Idaho. Page 22 General Technical Report INT-239. USDA Forest Service, Intermountain Research Station., Ogden, UT.
Noy-Meir, I. 1973. Desert ecosystems: environment and producers. Annual Review of Ecology and Systematics 4:25-51.
O’Connor, C., Miller, R. and Bates, J.D., 2013. Vegetation response to western juniper slash treatments. Environmental Management, 52(3): 553-566.
Olsen, R. 1992. Mule deer habitat requirements and management in Wyoming. B-965. University of Wyoming, Cooperative Extension Service, Laramie, WY.
Pellant, M. and C. Hall. 1994. Distribution of two exotic grasses in intermountain rangelands: status in 1992, USDA Forest Service Gen. Tech Report INT-GTR-313S: 109-112.
Pierson, F. B., C. J. Williams, P. R. Kormos, S. P. Hardegree, P. E. Clark, and B. M. Rau. 2010. Hydrologic vulnerability of sagebrush steppe following pinyon and juniper encroachment. Rangeland Ecology & Management 63:614-629.
Reynolds, T. D., Fraley., L. 1989. Root profiles of some native and exotic plant species in southeastern Idaho. Environmental and Experimental Botany 29(2): 241-248.
Richards, J. H. and M. M. Caldwell. 1987. Hydraulic lift: Substantial nocturnal water transport between soil layers by Artemisia tridentata roots. Oecologia 73:486-489.
Rickard, W. H., D. W. Uresk, and J. F. Cline. 1975. Impact of cattle grazing on three perennial grasses in south-central Washington. Journal of Range Management 28:108-112.
Ross, C. 1999. Population dynamics and changes in Curlleaf Mountain Mahogany in two adjacent Sierran and Great Basin mountain ranges. Ph.D. Dissertation, University of Nevada, Reno.
Russell, S.K. & Schupp, E.W. 1998. Effects of microhabitat patchiness on patterns of seed dispersal and seed predation of Cercocarpus ledifolius (Rosaceae). Oikos, 81, 434–443.
Sabo, J. L., et al. (2005). "Riparian zones increase regional species richness by harboring different, not more, species." Ecology 86(1): 56-62.
Sampson, A. W. and B. S. Jespersen (1963). California range brushlands and browse plants. Berkeley, CA, University of California, Division of Agricultural Sciences, California Agricultural Experiment Station, Extension Service: 162 p.
Sapsis, D. B. and J. B. Kauffman. 1991. Fuel consumption and fire behavior associated with prescribed fires in sagebrush ecosystems. Northwest Science 65:173-179.
Scheldt, R. S. 1969. Ecology and utilization of curl-leaf mountain mahogany in Idaho. Unpublished thesis. Moscow, ID, University of Idaho.
Schultz, B. W. 1987. Ecology of curl-leaf mountain mahogany (Cercocarpus ledifolius) in western and central Nevada: Population structure and dynamics. Master's thesis, University of Nevada, Reno. 111 pp.
Schultz, B. W., Tausch, R. J., Tueller, P.T. 1996. Spatial relationships among young Cercocarpus ledifolius (curlleaf mountain mahogany). Great Basin Naturalist 56(3): 261-266.
Schultz, B. W., Tausch, R., Tueller, P. T. 1991. Size, age, and density relationships to curlleaf mahogany (Cercocarpus ledifolus) populations in western and central Nevada: competitive implications. Western North American Naturalist 51:183-191.
Schultz, B. W., Tueller, P. T., Tausch, R. J. 1990. Ecology of curlleaf mahogany in western and central Nevada: community and population structure. Journal of Range Management 43: 13-20.
Sheley, R. L., E. A. Vasquez, A. Chamberlain, and B. S. Smith. 2012. Landscape-scale rehabilitation of medusahead (Taeniatherum caput-medusae)-dominated sagebrush steppe. Invasive Plant Science and Management 5(4):436-442.
Shumar, M. L., Anderson, J. E. 1986. Gradient analysis of vegetation dominated by two subspecies of big sagebrush. Journal of Range Management 39: 156-160.
Steele, R., R. D. Pfister, R. A. Ryker, and J. A. Kittams. 1981. Forest habitat types of central Idaho. Gen. Tech. Rep. INT-114. U. S. Department of Agriculture, Forest Service, Intermountain Research Station.
Stubbendieck, J. L., S. L. Hatch, and C. H. Butterfield. 1992. North American range plants. University of Nebraska Press, Lincoln, NE. 493 p.
Taylor, N. 2004. Distribution of domestic cattle in southern Utah vegetation communities, and, Predicting intake by elk and mule deer using blood and fecal samples. Utah State University.
Tueller, P. T. 1989. Vegetation and land use in Nevada. Rangelands 11:204-210.
Turner, G. T. 1969. Responses of mountain grassland vegetation to gopher control, reduced grazing, and herbicide. Journal of Range Management 22:377-383.
Uresk, D. W., and W. H. Rickard. 1976. Diets of steers on a shrub-steppe rangeland in south-central Washington. Journal of Range Management 29:464-466.
USDA Forest Service. 1937. Range Plant Handbook. New York, NY, Dover Publications.
USDA Forest Service. 1988. Range Plant Handbook. Dover Publications, Inc., New York, N. Y.
Vollmer, J. L. and J. G. Vollmer (2008). Controlling cheatgrass in winter range to restore habitat and endemic fire Proceedings-Shrublands under fire: disturbance and recovery in a changing world. RMRS-P-52., Cedar City, UT, Fort Collins, CO: US Department of Agriculture, Forest Service, Rocky Mountain Research Station.
Weisberg, P. J., T. E. Dilts, O. W. Baughman, S. E. Meyer, E. A. Leger, K. J. Van Gunst, and L. Cleeves. 2017. Development of remote sensing indicators for mapping episodic die-off of an invasive annual grass (Bromus tectorum) from the Landsat archive. Ecological Indicators 79:173-181.
Wood, M. K., B. A. Buchanan, and W. Skeet. 1995. Shrub preference and utilization by big game on New Mexico reclaimed mine land. Journal of Range Management 48:431-437.
Wright, H. A. 1971. Why squirreltail is more tolerant to burning than needle-and-thread. Journal of Range Management 24(4): 277-284.
Young, R. P. 1983. Fire as a vegetation management tool in rangelands of the intermountain region. Managing Intermountain rangelands - improvement of range and wildlife habitats: proceedings; 1981 September 15-17; Twin Falls, ID; 1982 June 22-24; Elko, NV Gen. Tech. Rep. INT-GTR-157., U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. Ogden, UT. 194 p.
Youngberg, C. T., and L. Hu. 1972. Root nodules on mountain mahogany. Forest Science 18(3):211-212.
Youngberg, C. T., Hu, L. 1972. Root nodules on mountain mahogany. Forest Science 18: 211-212.
Ziegenhagen, L. L. 2003. Shrub reestablishment following fire in the mountain big sagebrush (Artemisia tridentata Nutt. ssp. vaseyana (Rydb.) Beetle) alliance. M.s. Oregon State University.
Ziegenhagen, L. L., and R. F. Miller. 2009. Postfire recovery of two shrubs in the interiors of large burns in the Intermountain West, USA. Western North American Naturalist 69(2):195-205.
Contributors
DK/FR/SCB/GKB
Tamzen Stringham
Patti Novak-Echenique
Approval
Kendra Moseley, 4/10/2024
Rangeland health reference sheet
Interpreting Indicators of Rangeland Health is a qualitative assessment protocol used to determine ecosystem condition based on benchmark characteristics described in the Reference Sheet. A suite of 17 (or more) indicators are typically considered in an assessment. The ecological site(s) representative of an assessment location must be known prior to applying the protocol and must be verified based on soils and climate. Current plant community cannot be used to identify the ecological site.
| Author(s)/participant(s) | |
|---|---|
| Contact for lead author | |
| Date | 04/11/2024 |
| Approved by | Kendra Moseley |
| Approval date | |
| Composition (Indicators 10 and 12) based on | Annual Production |
Indicators
-
Number and extent of rills:
-
Presence of water flow patterns:
-
Number and height of erosional pedestals or terracettes:
-
Bare ground from Ecological Site Description or other studies (rock, litter, lichen, moss, plant canopy are not bare ground):
-
Number of gullies and erosion associated with gullies:
-
Extent of wind scoured, blowouts and/or depositional areas:
-
Amount of litter movement (describe size and distance expected to travel):
-
Soil surface (top few mm) resistance to erosion (stability values are averages - most sites will show a range of values):
-
Soil surface structure and SOM content (include type of structure and A-horizon color and thickness):
-
Effect of community phase composition (relative proportion of different functional groups) and spatial distribution on infiltration and runoff:
-
Presence and thickness of compaction layer (usually none; describe soil profile features which may be mistaken for compaction on this site):
-
Functional/Structural Groups (list in order of descending dominance by above-ground annual-production or live foliar cover using symbols: >>, >, = to indicate much greater than, greater than, and equal to):
Dominant:
Sub-dominant:
Other:
Additional:
-
Amount of plant mortality and decadence (include which functional groups are expected to show mortality or decadence):
-
Average percent litter cover (%) and depth ( in):
-
Expected annual annual-production (this is TOTAL above-ground annual-production, not just forage annual-production):
-
Potential invasive (including noxious) species (native and non-native). List species which BOTH characterize degraded states and have the potential to become a dominant or co-dominant species on the ecological site if their future establishment and growth is not actively controlled by management interventions. Species that become dominant for only one to several years (e.g., short-term response to drought or wildfire) are not invasive plants. Note that unlike other indicators, we are describing what is NOT expected in the reference state for the ecological site:
-
Perennial plant reproductive capability:
Print Options
Sections
Font
Other
The Ecosystem Dynamics Interpretive Tool is an information system framework developed by the USDA-ARS Jornada Experimental Range, USDA Natural Resources Conservation Service, and New Mexico State University.
Click on box and path labels to scroll to the respective text.