Natural Resources
Conservation Service
Ecological site R026XY038NV
LOAMY SLOPE 14+ P.Z.
Last updated: 4/10/2024
Accessed: 03/24/2026
General information
Provisional. A provisional ecological site description has undergone quality control and quality assurance review. It contains a working state and transition model and enough information to identify the ecological site.
MLRA notes
Major Land Resource Area (MLRA): 026X–Carson Basin and Mountains
The area lies within western Nevada and eastern California, with about 69 percent being within Nevada, and 31 percent being within California. Almost all this area is in the Great Basin Section of the Basin and Range Province of the Intermontane Plateaus. Isolated north-south trending mountain ranges are separated by aggraded desert plains. The mountains are uplifted fault blocks with steep side slopes. Most of the valleys are drained by three major rivers flowing east across this MLRA. A narrow strip along the western border of the area is in the Sierra Nevada Section of the Cascade-Sierra Mountains Province of the Pacific Mountain System. The Sierra Nevada Mountains are primarily a large fault block that has been uplifted with a dominant tilt to the west. This structure leaves an impressive wall of mountains directly west of this area. This helps create a rain shadow affect to MLRA 26. Parts of this eastern face, but mostly just the foothills, mark the western boundary of this area. Elevations range from about 3,806 feet (1,160 meters) on the west shore of Pyramid Lake to 11,653 feet (3,552 meters) on the summit of Mount Patterson in the Sweetwater Mountains.
Valley areas are dominantly composed of Quaternary alluvial deposits with Quaternary playa or alluvial flat deposits often occupying the lowest valley bottoms in the internally drained valleys, and river deposited alluvium being dominant in externally drained valleys. Hills and mountains are dominantly Tertiary andesitic flows, breccias, ash flow tuffs, rhyolite tuffs or granodioritic rocks. Quaternary basalt flows are present in lesser amounts, and Jurassic and Triassic limestone and shale, and Precambrian limestone and dolomite are also present in very limited amounts. Also of limited extent are glacial till deposits along the east flank of the Sierra Nevada Mountains, the result of alpine glaciation.
The average annual precipitation in this area is 5 to 36 inches (125 to 915 millimeters), increasing with elevation. Most of the rainfall occurs as high-intensity, convective storms in spring and autumn. Precipitation is mostly snow in winter. Summers are dry. The average annual temperature is 37 to 54 degrees F (3 to 12 degrees C). The freeze-free period averages 115 days and ranges from 40 to 195 days, decreasing in length with elevation.
The dominant soil orders in this MLRA are Aridisols and Mollisols. The soils in the area dominantly have a mesic soil temperature regime, an aridic or xeric soil moisture regime, and mixed or smectitic mineralogy. They generally are well drained, are clayey or loamy and commonly skeletal, and are very shallow to moderately deep.
This area supports shrub-grass vegetation characterized by big sagebrush. Low sagebrush and Lahontan sagebrush occur on some soils. Antelope bitterbrush, squirreltail, desert needlegrass, Thurber needlegrass, and Indian ricegrass are important associated plants. Green ephedra, Sandberg bluegrass, Anderson peachbrush, and several forb species also are common. Juniper-pinyon woodland is typical on mountain slopes. Jeffrey pine, lodgepole pine, white fir, and manzanita grow on the highest mountain slopes. Shadscale is the typical plant in the drier parts of the area. Sedges, rushes, and moisture-loving grasses grow on the wettest parts of the wet flood plains and terraces. Basin wildrye, alkali sacaton, saltgrass, buffaloberry, black greasewood, and rubber rabbitbrush grow on the drier sites that have a high concentration of salts.
Some of the major wildlife species in this area are mule deer, coyote, beaver, muskrat, jackrabbit, cottontail, raptors, pheasant, chukar, blue grouse, mountain quail, and mourning dove. The species of fish in the area include trout and catfish. The Lahontan cutthroat trout in the Truckee River is a threatened and endangered species.
LRU notes
The Sierra Influenced Ranges LRU is characterized by wooded great basin mountains with climatic and biotic affinities to the Sierra Nevada mountain range. The Sierra Influences Ranges LRU receives greater precipitation that the mountain ranges of central NV. Amount of precipitation varies in relation to the local strength of the Sierra NV rain shadow, characterized by pinyon and juniper trees. The White, Sweetwater, Pine Nut, Wassuk, and Virginia ranges of Nevada support varying amounts of Sierra Nevada flora, such as ponderosa pine. Elevations range from 1610 to 2420 meters and slopes range from 5 to 49 percent, with a median value of 22 percent. Frost free days (FFD) ranges from 92 to 163.
Ecological site concept
The Loamy Slope 14+ P.Z. site occurs on smooth to slightly concave mountain sideslopes. Although this community occurs on all aspects, it is usually restricted to northerly aspects at lower elevations. Slopes range from 15 to 75 percent. Elevations are 4500 to 9500. The soils are typically more than 40 inches deep and well drained. The soils have formed in residuum and colluvium from intermediate volcanic and granitic parent materials. The dominant plants are mountain big sagebrush (Artemisia tridentata ssp. vaseyana) and western needlegrass (Achnatherum occidentale).
Associated sites
| R026XY028NV |
MOUNTAIN RIDGE |
|---|---|
| R026XY039NV |
CLAYPAN 14+ P.Z. |
| R026XY084NV |
DEEP LOAMY 14+ P.Z. |
Similar sites
| R026XY018NV |
GRANITIC SOUTH SLOPE 10-12 P.Z. ACTH7-ACOCO codominant |
|---|---|
| R026XY100NV |
STONY SLOPE 10-12 P.Z. ACTH7-POFE codominant |
| R026XY082NV |
MOUNTAIN LOAM 16+ P.Z. ACPI2-KOMA codominant grasses with ACLE9 |
| R026XY076NV |
MOUNTAIN SHOULDERS 16+ P.Z. ACLE9 dominant grass; less productive site |
| R026XY089NV |
SOUTH SLOPE 12-14 P.Z. LEKI2-ACHNA codominant |
| R026XY079NV |
GRANITIC SOUTH SLOPE 14+ P.Z. Soils derived from granitic parent materials |
| R026XY053NV |
LOAMY 16+ P.Z. BRMA4 codominant grass |
| R026XY048NV |
LOAMY SLOPE 12-14 P.Z. PUTR2 codominant shrub; ACTH7 codominant grass; more productive site |
| R026XY008NV |
GRANITIC FAN 10-12 P.Z. HECO26-ACHY codominant grasses |
| R026XY046NV |
GRANITIC SLOPE 12-14 P.Z. less productive site |
| R026XY026NV |
GRANITIC SLOPE 10-12 P.Z. ACTH7-ACSP12 codominant |
| R026XY005NV |
LOAMY 12-14 P.Z. More productive site |
| R026XY052NV |
SHALLOW LOAM 16+ P.Z. LEKI2 dominant grass |
| R026XY040NV |
GRAVELLY LOAM 14+ P.Z. PUTR2 dominant shrub; more productive site |
| R026XY084NV |
DEEP LOAMY 14+ P.Z. ACOCO-PONE3 codominant |
| R026XY010NV |
LOAMY 10-12 P.Z. ACTH7 dominant grass |
| R026XY006NV |
GRANITIC LOAM 14+ P.Z. Less productive site; soils derived from granitic parent materials |
Table 1. Dominant plant species
| Tree |
Not specified |
|---|---|
| Shrub |
(1) Artemisia tridentata ssp. vaseyana |
| Herbaceous |
(1) Achnatherum occidentale |
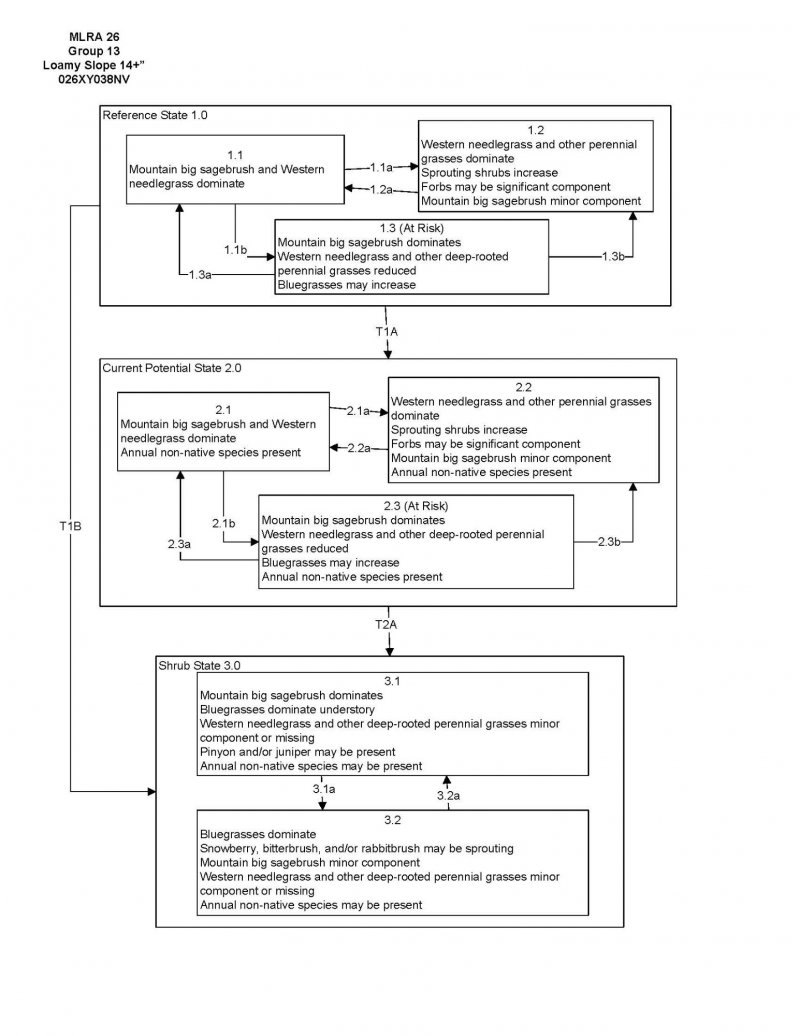

Click on box and path labels to scroll to the respective text.