
Natural Resources
Conservation Service
Ecological site R030XB166CA
Dissected Pediment, Cool
Last updated: 10/21/2024
Accessed: 04/10/2026
General information
Provisional. A provisional ecological site description has undergone quality control and quality assurance review. It contains a working state and transition model and enough information to identify the ecological site.
Figure 1. Mapped extent
Areas shown in blue indicate the maximum mapped extent of this ecological site. Other ecological sites likely occur within the highlighted areas. It is also possible for this ecological site to occur outside of highlighted areas if detailed soil survey has not been completed or recently updated.
MLRA notes
Major Land Resource Area (MLRA): 030X–Mojave Basin and Range
MLRA Description:
Major Land Resource Area (MLRA) 30, Mojave Desert, is found in southern California, southern Nevada, the extreme southwest corner of Utah and northwestern Arizona within the Basin and Range Province of the Intermontane Plateaus. The climate of the area is hot (primarily hyperthermic and thermic; however at higher elevations, generally above 5000 feet, mesic, cryic and frigid) and dry (aridic). Elevations range from below sea level to over 12,000 feet in the higher mountain areas found within the MLRA. Due to the extreme elevational range found within this MLRA, Land Resource Units (LRUs) were designated to group the MLRA into similar land units.
LRU Description:
This LRU (designated by ’XB’) is found across the eastern half of California, much of the mid-elevations of Nevada, the southernmost portions of western Utah, and the mid-elevations of northwestern Arizona. Elevations range from 1800 to 5000 feet and precipitation ranges from 4 to 9 inches per year, but is generally between 5-6 inches. This LRU is characterized primarily by the summer precipitation it receives, ranging from 18 – 35% but averages 25%. Summer precipitation falls between July and September in the form of rain, and winter precipitation falls starting in November and ends between February and March, also mostly in the form of rain; however it does receive between 0 and 3 inches of snow, with an average of 1 inch. The soil temperature regime is thermic and the soil moisture regime is typic-aridic. Vegetation includes creosote bush, burrobush, Nevada jointfir, ratany, Mojave yucca, Joshua tree, chollas, cactus, big galleta grass and several other warm season grasses. At the upper portions of the LRU, plant production and diversity are greater and blackbrush is a common dominant shrub.
Ecological site concept
This ecological site occurs on granitic pediments and more stable areas of fan remnants on top of granitic pediments at elevations of approximately 3300 to 5200 feet. Soils are typically very shallow to shallow loamy sands with high run-off. Production reference value (RV) is 249 pounds per acre and ranges from 92 to 438 pounds per acre depending on annual precipitation and annual species production. Blackbrush (Coleogyne ramosissima) and California juniper (Juniperus californica) dominate the plant community, and are supported by the cool thermic climate. Shallow soils support blackbrush as a dominant species. The dissected pediment landscape with localized run-off and run-on supports a high diversity of shrubs relative to the strongly blackbrush stands found on more homogenous landscapes.
Data ranges in the physiographic data, climate data, water features, and soil data sections of this Ecological Site Description are based on major components only (15 percent of mapunit or greater).
This site is is part of grouped provisional concept R030XB076NV.
Associated sites
| R030XB168CA |
Cool Deep Sandy Fans This ecological site is found on adjacent fan aprons. The dominant species are blackbrush (Coleogyne ramosissima) and California juniper (Juniperus californica). |
|---|---|
| R030XB173CA |
Coarse Loamy Very Deep Fan Remnants This ecological site is found on adjacent fan remnants. Blackbrush (Coleogyne ramosissima), Joshua tree (Yucca brevifolia) and big galleta (Pleuraphis rigida) are dominant species. |
| R030XB174CA |
Sandy Fan Aprons This ecological site is found on adjacent fan aprons. Creosote bush (Larrea tridentata), Joshua tree (Yucca brevifolia) and big galleta (Pleuraphis rigida) are dominant species. |
| R030XB189CA |
Shallow Cool Hills This ecological site is found on adjacent hills and mountains. The dominant species are blackbrush (Coleogyne ramosissima) and California juniper (Juniperus californica). |
| R030XY202CA |
Very Rarely To Rarely Flooded Thermic Ephemeral Stream This ecological site is found on adjacent very rarely flooded ephemeral drainageways. California jointfir (Ephedra californica), Mojave indigobush (Psorothamnus arborescens) and burrobrush (Hymenoclea salsola) are dominant species. |
Similar sites
| R030XB189CA |
Shallow Cool Hills R030XB189 is found on hills and mountains. Plant cover, density and production are typically higher. |
|---|---|
| R030XB225CA |
Warm Sloping Pediments R030XB225CA occurs on lower elevation pediments, and is co-dominated by burrobush (Ambrosia dumosa) and hall’s shrubby spurge (Tetracoccus hallii). Blackbrush (Coleogyne ramosissima) is only trace if present. |
| R030XB228CA |
Warm Shallow Pediments R030XB228CA occurs on pediments at lower elevations, and is co-dominated by creosote bush (Larrea tridentata) and burrobush (Ambrosia dumosa). Blackbrush (Coleogyne ramosissima) is not present. |
| R030XB188CA |
Cool Shallow to Moderately Deep Fans R030XB188CA is found on pediments overlain with a thicker layer of alluvium. Creosote bush (Larrea tridentata) is co-dominant with blackbrush (Coleogyne ramosissima). |
| R030XB221CA |
Loamy Fan Remnants And Pediments R030XB221CA occurs on pediments and fan remnants with moderately deep soils to a duripan. It is dominated by blackbrush (Coleogyne ramosissima), burrobush (Ambrosia dumosa) and hall’s shrubby spurge (Tetracoccus hallii). |
| R030XB171CA |
Dissected Pediment R030XB171CA occurs on pediments at lower elevations. Blackbrush (Coleogyne ramosissima) is weakly dominant with a high diversity of other shrubs, including burrobush (Ambrosia dumosa) and creosote bush (Larrea tridentata). California juniper (Juniperus californica) is not present. |
Table 1. Dominant plant species
| Tree |
(1) Juniperus californica |
|---|---|
| Shrub |
(1) Coleogyne ramosissima |
| Herbaceous |
Not specified |
Physiographic features
This ecological site is found on pediments at elevations of 3280 to 5180 feet. Slopes range from 4 to 30 percent. This site experiences no flooding or ponding, and runoff class is high.
Table 2. Representative physiographic features
| Landforms |
(1)
Pediment
|
|---|---|
| Flooding frequency | None |
| Ponding frequency | None |
| Elevation | 3,280 – 5,180 ft |
| Slope | 4 – 30% |
| Aspect | Aspect is not a significant factor |
Climatic features
The climate on this site is arid characterized by cool, somewhat moist winters and hot, dry summers. The average annual precipitation ranges from 4 to 7 inches with most falling as rain from November to March with a secondary, smaller peak between July and September occurring as summer convection storms. Mean annual air temperature ranges from 55 to 63 degrees F. Average maximum temperatures in the summer months range from 85 to 90 degrees F, and average minimum temperatures in the winter months range from 37 to 40 degrees F. The frost free period is 210 to 280 days.
Maximum and minimum monthly climate data for this ESD were generated by the Climate Summarizer
(http://www.nm.nrcs.usda.gov/technical/handbooks/nrph/Climate_Summarizer.xls) using data
from the following climate stations (results are weighted averages; numbers in square brackets represent relative weights):
44405 JOSHUA TREE, CA (Period of record = 1959 to 2011) [1]
LTHC1 Lost Horse, Joshua Tree National Park
(Period of record = 1991 to 2011) [2]
44467 Kee Ranch, CA (Period of record = 1948 to 1979) [1]
The data from multiple weather were combined to most accurately reflect the climatic conditions of this ecological site. The Lost Horse weather station is closest to this ecological site but is limited by the number of years data was collected. The Joshua Tree weather station is also nearby this ecological site but is at slightly lower elevation, and is lacking precipitation data for the years between 1975 and 2008. The Kee Ranch weather station contains precipitation data for all years of the period of record but has no temperature data.
Table 3. Representative climatic features
| Frost-free period (average) | 280 days |
|---|---|
| Freeze-free period (average) | |
| Precipitation total (average) | 7 in |
Figure 2. Monthly precipitation range
Figure 3. Monthly average minimum and maximum temperature
Influencing water features
Soil features
The soils associated with this ecological site are very shallow to shallow, and well to somewhat excessively drained. These soils formed in alluvium or colluvium derived from granite or gneiss over residuum weathered from granite or gneiss. Surface textures are typically loamy sand, gravelly loamy sand, and sand and less commonly, sandy loam. Subsurface horizons are mainlysand, loamy sand, and loamy coarse sand, with sandy loam textures only found on areas of fan remnant over pediment. For rock fragments less than 3 inches in diameter, the percent surface cover ranges from 5 to 65 percent, and subsurface volume ranges from 1 to 20 percent (subsurface fragments by volume are RV ranges for all soil profile horizons for a depth of 0 to 14 inches). For rock fragments greater than 3 inches in diameter, the percent surface cover ranges from 4 to 20 percent, and subsurface volumes range from 0 to 10 percent.
This ecological site is associated with the following soil series: Pinecity (Mixed, thermic, shallow Typic Torripsamments), and Desertqueen (Loamy, mixed, superactive, thermic, shallow Typic Haplargids). Typically this site is found on Pinecity soils which have sandy textures directly above weathered granitic bedrock between 5 and 36 centimeters. Pinecity soils are commonly found on low sloping pediment, but can also occur on backslopes of low hills. This site is less commonly found on Desertqueen soils which have a sandy loam argillic horizon (3 to 20 centimeters deep) above weathered granite or gneiss (12 to 36 centimeters deep). Neither soil is skeletal (having >35% rock fragments) in the soil profile, and typically the soil surface is covered by gravel. Both soils are commonly found in map units with greater than 15 percent rock outcrop in the unit.
This ecological site is correlated with the following map units and soil components in the Joshua Tree National Park Soil Survey:
4607;Pinecity sand, 4 to 8 percent slopes;Pinecity;;85; Pinecity;sloping;5
4608;Pinecity-Rock outcrop association, 4 to 15 percent slopes, high elevation;Pinecity;;60
4606;Pinecity-Rock outcrop association, 4 to 15 percent slopes;Pinecity;;60; Desertqueen;cool;5
4615;Desertqueen-Jumborox-Rock outcrop association, 2 to 15 percent slopes;Desertqueen;cool;45; Pinecity;;5
3685;Morongo-Desertqueen complex, 8 to 30 percent slopes; Desertqueen; undulating;15; Pinecity;;2
3681;Morongo-Jumborox complex, 4 to 8 percent slopes, warm;Pinecity;;1
4071;Helendale-Desertqueen association, 4 to 15 percent slopes;Pinecity;;5
4091;Littlefargo-Rock outcrop, 4 to 15 percent slopes;Pinecity;;3
Table 4. Representative soil features
| Parent material |
(1)
Alluvium
–
granite
(2) Residuum – gneiss |
|---|---|
| Surface texture |
(1) Loamy sand (2) Sand (3) Sandy loam |
| Family particle size |
(1) Sandy |
| Drainage class | Well drained to somewhat excessively drained |
| Permeability class | Moderate to rapid |
| Soil depth | 2 – 14 in |
| Surface fragment cover <=3" | 5 – 65% |
| Surface fragment cover >3" | 1 – 20% |
| Available water capacity (0-40in) |
0.2 – 1.6 in |
| Calcium carbonate equivalent (0-40in) |
1% |
| Electrical conductivity (0-40in) |
2 mmhos/cm |
| Sodium adsorption ratio (0-40in) |
4 |
| Soil reaction (1:1 water) (0-40in) |
6.6 – 8.4 |
| Subsurface fragment volume <=3" (Depth not specified) |
4 – 20% |
| Subsurface fragment volume >3" (Depth not specified) |
10% |
Ecological dynamics
Abiotic Factors
A shallowly dissected pediment landform, shallow soils and a cool thermic climate are the most important abiotic factors driving this ecological site.
The cool thermic climate supports dominance by blackbrush and California juniper. At lower elevations, warmer, drier climates support co-dominance by creosote bush (Larrea tridentata) and burrobush (Ambrosia dumosa), and exclude California juniper. At higher elevations, moister climates support pinyon-juniper woodlands.
The majority of the landscape is characterized by shallow soils that support dominance by blackbrush. Blackbrush is a shallow-rooted, very long-lived, drought-tolerant shrub. Shallow roots enable blackbrush to thrive in soils where soil moisture is restricted to shallow depths, and extreme drought-tolerance coupled with extreme longevity allow blackbrush to achieve community dominance over shorter-lived, less drought-tolerant species, provided there are long periods of time without disturbance (Pendleton and Meyer 2004). California juniper is a long-lived evergreen shrub or small tree that is often associated with shallow soils (Sawyer et al. 2009). It achieves maximum dominance at elevations slightly higher than this ecological site, where mean annual precipitation is above 9.5 inches (Rhode 2002). Thus, while California juniper is an important species in this ecological site, it is restricted to mesic microsites receiving additional run-on, and does not achieve dominance across the site. Some secondary species that occur on this site are also restricted to areas receiving additional run-on, including desert almond (Prunus fasciculata) and catclaw acacia (Acacia gregii).
Pediments are gently sloping, bedrock-driven surfaces of low relief that form at the base of receding mountain fronts. Pediment surfaces are covered by a discontinuous layer of alluvium that is typically thinner closer to the mountain front and eventually thick enough that the pediment is no longer distinct from the alluvial fan (Dohrenwend and Parsons 2009). Closer to the mountain front erosion is most intense, and exposed bedrock is dissected and undulating. Along the fan piedmont part of the landscape, erosion is less active, the layer of alluvium is thicker, and the landscape is less dissected (Dohrenwend and Parsons 2009). This ecological site occurs in positions that do not receive intense sheetflow, are covered by a thin layer of alluvium, have patches of exposed bedrock and a shallowly dissected moderately undulating landscape.
Different topographical positions within these dissected pediment landscapes experience different rates of erosion and deposition (Abrahams and Parsons 1991, Edinger-Marshall and Lund 1999). The dissected pediment landscape, with contrasting and localized areas of soil erosion and deposition, and run-off and run-on, supports a relatively diverse shrub community. The low level disturbance caused by erosion supports the continuous presence of shorter-lived subshrubs. Annual species are generally limited on pediment landforms due to low soil moisture availability in shallow soils, and by soil erosion which limits seedling establishment (Guerrero-Campo et al. 2008), but at the higher, more mesic elevation range of this ecological site, annuals may be relatively abundant.
Disturbance dynamics
The major disturbances affecting this ecological site are drought, invasion by non-native species, fire and erosion.
Drought is an important shaping force in Mojave Desert plant communities (Webb et al. 2003, Hereford et al. 2006). Short-lived perennial shrubs and perennial grasses demonstrate the highest rates of mortality (Webb et al. 2003, Bowers 2005, Hereford et al. 2006, Miriti et al. 2007), and annual species remain dormant in the soil seedbank (Beatley 1969, 1974, 1976). Long-lived shrubs are more likely to exhibit branch-pruning, and or limited recruitment during drought (e.g. Hereford et al. 2006, Miriti et al. 2007), leading to reduced cover and biomass in drought-afflicted communities. Because relatively sparse cover characterizes this ecological site, further loss of cover due drought-induced mortality increases the susceptibility of this site to increased damages from erosion.
Non-native annual grasses (red brome [Bromus rubens], cheatgrass [Bromus tectorum] and Mediterranean grass [Schismus species]) have become naturalized throughout the Mojave Desert over the past century (Rickard and Beatley 1965, D'Antonio and Vitousek 1992, Brooks 1999, Reid et al. 2006, Norton et al. 2007). Increased soil nitrogen due to air pollution has been shown to increase the productivity of exotic annual grasses across the Mojave Desert (Brooks 1999). In Joshua Tree National Park, elevated soil N is decreasing the diversity and cover of native forb species in the park, and at the same time increasing the cover of exotic annual grasses (Rao and Allen 2010). Erosion and low soil moisture availability in the shallow soils of this site limits seedling establishment and productivity of annual species (Guerrero-Campo and Montserrat-Marti 2000). However, at the higher elevation range of this site where rainfall is higher, or in locations closer to urban centers receiving higher rates of nitrogen deposition, non-native grass abundance and productivity may be high.
Invasion by non-native annual grasses has increased the flammability of Mojave Desert vegetation communities by providing a continuous fine fuel layer between widely spaced shrubs (Brown and Minnich 1986, Brooks 1999, Brooks et al. 2004, Rao and Allen 2010 2010, Rao et al. 2010). After fire, these communities appear to be more susceptible to invasion by exotic grasses, leading to a grass-fire cycle (D'Antonio and Vitousek 1992). Historically, fire would have been a very rare occurrence in this ecological site due to the lack of a continuous fine fuel layer (Minnich 2003, Webb et al. 1987), occurring only in the year immediately following heavy winter precipitation or during periods of extreme fire behavior (Brooks et al. 2007).
Water erosion is the dominant process modifying and maintaining exposed pediment surfaces (Edinger-Marshall and Lund 1999, Dohrenwend and Parsons 2009), and is an important process modifying and maintaining the vegetation community. Without additional disturbance, the affects of erosion are within the natural range of variability of the reference plant community. However, with additional disturbance (anthropogenic or natural), the effects of erosion may be more severe, leading to vegetation and soil loss and potentially a new community phase or state. For example, shrub cover protects the soils of this ecological site from erosion (Edinger-Marshall and Lund 1999). If shrub cover is reduced due to drought or fire, erosion will remove more soil, which reduces the availability of safe sites for plant establishment, which further reduces shrub cover.
State and transition model
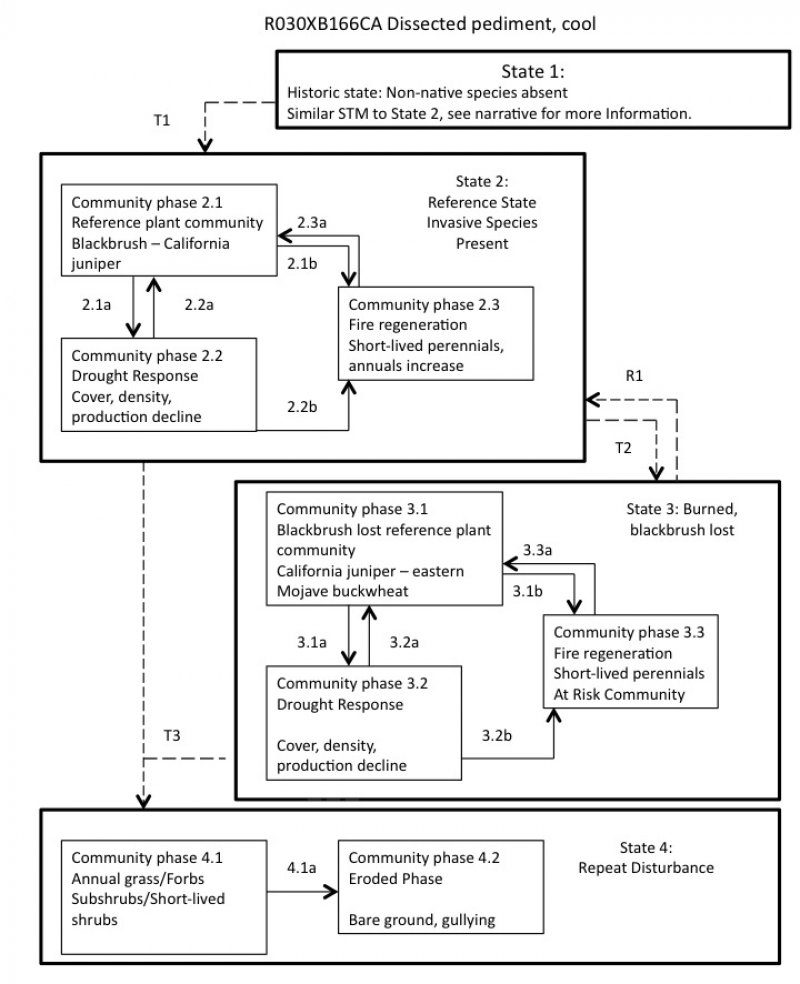
Figure 4. R030XB166CA
More interactive model formats are also available.
View Interactive Models
More interactive model formats are also available.
View Interactive Models
Click on state and transition labels to scroll to the respective text
State 2 submodel, plant communities
State 3 submodel, plant communities
State 4 submodel, plant communities
State 1
Historic State
State 1 represents the historic range of variability for this ecological site. This state no longer exists due to the ubiquitous naturalization of non-native species in the Mojave Desert. Periodic drought, erosion, and very rare fire were the natural disturbances influencing this ecological site. Blackbrush is killed by moderate to severe fire, and the historic fire return interval is suggested to be upwards of 100 years (Webb et al. 1987, Brooks et al. 2007, Abella et al. 2009). With the absence of non-native species to fuel recurrent fires, this long fire-return interval allowed for recovery to pre-burn densities. California juniper is also killed by moderate to severe fire, but in can re-colonize burned areas if the fire return interval is greater than 15 years (Cope 1992). Data for this State does not exist, but it would have been similar to State 2, except with only native species present. See State 2 narrative for more detailed information.
State 2
Reference State
State 2 represents the current range of variability for this site. Non-native annuals, including red brome and cheatgrass are naturalized in this plant community. Their abundance varies with precipitation, but they are at least sparsely present (as current year's growth or present in the soil seedbank).
Community 2.1
Reference plant community

Figure 5. Community Phase 2.1

Figure 6. Community Phase 2.1

Figure 7. Community Phase 2.1

Figure 8. Community Phase 2.1
The current potential plant community is dominated by blackbrush, with California juniper dispersed throughout the site in areas receiving additional run-on. Important secondary shrubs include Acton’s encelia (Encelia actonii), Nevada jointfir (Ephedra nevadensis), Mojave yucca (Yucca schidigera) and eastern Mojave buckwheat. A relatively high diversity of minor shrubs may also be present, and are often found near areas of additional run-on, such as at the base of rock outcrops, or in concave areas. Areas of run-on may support desert almond (Prunus fasciculata) and catclaw acacia (Acacia gregii). Subshrubs are an important component of the vegetation community, and include shrubby deervetch (Lotus rigidus), desert globemallow (Sphaeralcea ambigua), and whishbone bush (Mirabilis laevis var. villosa). The perennial bunchgrass desert needlegrass (Achnatherum speciosum) is usually sparsely present. Native winter annuals are seasonally present, as are the non-native annual grasses red brome and cheatgrass. Biological soil crusts (BSC) are often associated with the grussy granite (granite that is granulated but not decomposed) typical of soil surfaces on this ecological site. These crusts are important for improving soil stability, infiltration, and nutrient cycling on these shallow soils (Belnap et al. 2001). Biological soil crusts form slowly, and are very sensitive to physical disturbance (such as from trampling or off-road vehicle disturbance).
Figure 9. Annual production by plant type (representative values) or group (midpoint values)
Table 5. Annual production by plant type
| Plant type | Low (lb/acre) |
Representative value (lb/acre) |
High (lb/acre) |
|---|---|---|---|
| Shrub/Vine | 80 | 160 | 240 |
| Grass/Grasslike | 3 | 9 | 85 |
| Tree | 5 | 40 | 63 |
| Forb | 4 | 40 | 50 |
| Total | 92 | 249 | 438 |
Table 6. Canopy structure (% cover)
| Height Above Ground (ft) | Tree | Shrub/Vine | Grass/ Grasslike |
Forb |
|---|---|---|---|---|
| <0.5 | – | 1-2% | 1-3% | 1-3% |
| >0.5 <= 1 | – | 15-25% | 1-3% | 1-3% |
| >1 <= 2 | – | 1-5% | – | 0-1% |
| >2 <= 4.5 | – | 1-5% | – | – |
| >4.5 <= 13 | – | – | – | – |
| >13 <= 40 | – | – | – | – |
| >40 <= 80 | – | – | – | – |
| >80 <= 120 | – | – | – | – |
| >120 | – | – | – | – |
Community 2.2
Drought Response
This community phase is characterized by a decline in cover and production due to branch-pruning of long-lived shrubs (including blackbrush, Nevada jointfir, and Mojave yucca), mortality of California juniper, and mortality of shorter-lived perennials (including Acton’s brittlebush, eastern Mojave buckwheat, and desert needlegrass), and lack of emergence of annual forbs and grasses. This is an at-risk phase, as the increase in bare ground that occurs during drought increases the susceptibility of this site to erosion. Biological soil crusts are dormant during drought, and are especially susceptible to damage by mechanical disturbance when dry (Warren and Eldridge 2003). Thus, any additional disturbance threatens to transition this community phase to a phase of increased erosion, or a new state, where significant loss of ecological function has occurred.
Community 2.3
Fire regeneration community
This community phase is characterized by high species evenness following the loss of blackbrush and California juniper. Initially the post-burn community is dominated by non-native annual grasses (red brome and cheatgrass), annual forbs (including desert dandelion [Malacothrix glabrata], bristly fiddleneck [Amsinckia tessellata] and pincushion flower [Chaenactis fremontii]), subshrubs (including desert globemallow, wishbone bush and shrubby deervetch) and perennial grasses (desert needlegrass, Indian rice grass [Achnatherum hymenoide]). With time, species capable of resprouting after fire become more important, including Mojave yucca, Nevada ephedra, catclaw acacia, and desert almond. Species capable of quickly colonizing after fire include eastern Mojave buckwheat, turpentine bush, Cooper’s goldenbush, narrowleaf goldenbush, and burrobrush (Hymenoclea salsola). As tall shrub cover increases, shade-dependent seedlings of California juniper begin to establish. With very long periods of time with no disturbance, blackbrush re-colonizes very gradually replaces by shorter-lived species (Vasek 1983, Abella 2009, Vamstad 2009). This community is an at-risk phase. The increased cover and biomass of non-native annual grasses increases the likelihood of repeat burning (Brooks et al. 2004, Brooks and Matchett 2006, Brooks et al. 2007). If the fire return interval is less than 100 years, this community is very likely to transition to State 3. Reduction in shrub cover increases bare ground and increases the risk of erosion.
Pathway 2.1a
Community 2.1 to 2.2
This pathway occurs with severe or prolonged drought.
Pathway 2.1b
Community 2.1 to 2.3
This pathway occurs with moderate to severe fire.
Pathway 2.2a
Community 2.2 to 2.1
This pathway occurs with a return to average climatic conditions. Growth of long-lived shrubs and colonization by shorter-lived shrubs increases cover.
Pathway 2.2b
Community 2.2 to 2.3
This pathway occurs with moderate to severe fire. Although live annuals are largely absent from Community Phase 2.2, standing annual biomass in drought years immediately following a period of heavy precipitation poses a severe risk for fire. Cured native annual cover may pose a risk during the first year of drought, and non-native annual grasses pose a risk for three or more years (Minnich 2003, Brooks et al. 2007, Rao et al. 2010).
Pathway 2.3a
Community 2.3 to 2.1
This pathway occurs with a long period of time without disturbance (> 100 years).
State 3
Burned, blackbrush lost
This state is characterized by the loss of blackbrush from the plant community due to severe or recurrent fire.
Community 3.1
Blackbrush lost Reference plant community

Figure 10. Community Phase 3.1
This community represents the post-fire potential plant community for this site. This community is dominated by eastern Mojave buckwheat, with California juniper dispersed throughout the site in areas receiving additional runon. Important secondary shrubs include Mojave yucca, Nevada ephedra, and Cooper’s goldenbush. The perennial bunchgrass desert needlegrass is typically sparsely present, and big galleta may be present. Subshrubs are an important component of the plant community, and include shrubby deervetch, wishbone bush and desert globemallow. Winter annuals may be seasonally abundant, generally at higher levels than in the Reference State. The non-native annual grasses red brome and cheatgrass are present, as is the non-native annual forb red-stem stork’s bill.
Figure 11. Annual production by plant type (representative values) or group (midpoint values)
Table 7. Annual production by plant type
| Plant type | Low (lb/acre) |
Representative value (lb/acre) |
High (lb/acre) |
|---|---|---|---|
| Shrub/Vine | 150 | 225 | 300 |
| Forb | 25 | 75 | 100 |
| Grass/Grasslike | 20 | 35 | 25 |
| Tree | 12 | 20 | 25 |
| Total | 207 | 355 | 450 |
Community 3.2
Drought Response
This community phase is characterized by a decline in cover and production due to branch-pruning of long-lived shrubs including Nevada ephedra and Mojave yucca, mortality of California juniper, and mortality of shorter-lived perennials (including eastern Mojave buckwheat, burrobrush, Cooper’s goldenbush, and desert needlegrass), and lack of emergence of annual forbs and grasses. See narrative for Community Phase 2.2 for more information.
Community 3.3
Fire regeneration community
This community phase develops after a moderate to severe burn. California juniper is largely lost from the community due to fire-induced mortality, and this community phase is characterized by high species evenness. Initially the post-burn community is dominated by non-native annual grasses, annual forbs, subshrubs, and perennial grasses. With time, species capable of resprouting after fire become more important, including Mojave yucca, Nevada ephedra, catclaw acacia, and desert almond. Species capable of quickly colonizing after fire include eastern Mojave buckwheat, turpentine bush, Cooper’s goldenbush, narrowleaf goldenbush, and burrobrush. As tall shrub cover increases, shade-dependent seedlings of California juniper begin to establish, and California juniper gradually increases in abundance. This community is an at-risk phase. The increased cover and biomass of non-native annual grasses increases the likelihood of repeat burning (Brooks et al. 2004, Brooks and Matchett 2006, Brooks et al. 2007). If the fire return interval is less than 100 years, this community is very likely to transition to State 3. Reduction in shrub cover increases bare ground and increases the risk of erosion.
Pathway 3.1a
Community 3.1 to 3.2
This pathway occurs in response to prolonged or severe drought.
Pathway 3.1b
Community 3.1 to 3.3
This pathway occurs in response to moderate or severe fire.
Pathway 3.2a
Community 3.2 to 3.1
This pathway occurs with a return to average or above average precipitation.
Pathway 3.2b
Community 3.2 to 3.3
This pathway occurs with fire. Although live annuals are largely absent from Community Phase 3.2, standing annual biomass in drought years immediately following a period of heavy precipitation poses a severe risk for fire. Cured native annual cover may pose a risk during the first year of drought, and non-native annual grasses pose a risk for three or more years (Minnich 2003, Brooks et al. 2007, Rao et al. 2010).
Pathway 3.3a
Community 3.3 to 3.1
This pathway occurs with time without fire.
State 4
Repeat Disturbance
This state develops after repeat disturbance in the reference or blackbrush lost communities. This state has been significantly altered from the natural range of variability found in States 1 through 3. Blackbrush and California juniper have been lost, and non-native annual grasses, native sub-shrubs, perennial grasses, and short-lived shrubs dominate the community, or in the eroded phase, bare ground dominates.
Community 4.1
Repeated Fire
This community phase occurs when the fire return interval is less than 15 years. The plant community is dominated by non-native annual species including red brome, cheatgrass, Mediterranean grass and red-stem stork’s bill, and native forbs, including desert dandelion, bristly fiddleneck and pincushion flower (many other native forbs could also be present). Native subshrubs including desert globemallow, wishbone bush and shrubby deervetch may be abundant, as might the perennial bunchgrass desert needlegrass. There may be very sparse cover of resprouting shrubs, but with recurring fire, these are likely to die out. Short-lived shrubs may be sparsely present, and may include Cooper’s goldenbush, burrobrush, and eastern Mojave buckwheat. This community is at high-risk of repeat burning due to high fine fuel cover. This community is also susceptible to wind and water erosion, due to the loss of stabilizing shrub cover (Bull 1997). This can lead to gullying, loss of important topsoil, and exposure of subsurface horizons that are not conducive to plant establishment, and further degrade the site. Furthermore, the loss of vegetation structure present in States 2 and 3 reduces the suitability of this habitat for wildlife (Brooks et al. 2007, Vamstad 2009). Since rodent seed-caching is important for the dispersal and establishment of many desert species, this can further inhibit recovery.
Community 4.2
Eroded Phase
This community phase is characterized by severe soil erosion. Biological soil crusts are largely absent, gullying is pronounced, and soil surfaces have no protective surface gravels. Bare ground dominates the landscape, with vegetative cover restricted to interfluves or sheltered locations around rock outcrops. Shrubs capable of resprouting or of quickly colonizing after disturbance increase in importance, including subshrubs (see Community phase 2.1 list), and catclaw acacia, Acton’s encelia, burrobrush, Coopers’ goldenbush and eastern Mojave buckwheat. Blackbrush is absent or only trace.
Pathway 4.1a
Community 4.1 to 4.2
This pathway occurs with additional disturbance such as drought followed by severe rain or wind, or anthropogenic disturbance such as trampling and off-road vehicle use.
Transition 1
State 1 to 2
This transition occurred with the naturalization of non-native species in this ecological site. Non-native species were introduced with settlement of the Mojave Desert region in the 1860s.
Transition 2
State 2 to 3
This transition occurs with extensive, severe fire when blackbrush seed sources are not available to colonize burned areas, or with recurrent fire.
Restoration pathway 1
State 3 to 2
Restoration of arid desert communities severely altered by repeat fire at the landscape scale is very difficult (Allen 1993). Reducing invasion of non-native grasses that increase after fire may help promote native plant recovery, and reduce the probability of repeat burning (Fuhrmann et al. 2009, Matchett et al. 2009, Steers and Allen 2010); however, accomplishing this at a landscape scale, for a time period long enough to be effective, has not yet been accomplished. In small-scale trials, Fusilade, a grass-specific herbicide, was successful in reducing invasive grasses in burned creosote bush communities in the Colorado Desert in the initial three years after fire (Steers and Allen 2010). The long-term efficacy of such treatments on a landscape scale, and non-target effects have not yet been determined. The pre-emergent herbicide Plateau was applied in conjunction with aerial seeding of natives after fire in Zion National Park (Fuhrmann et al. 2009, Matchett et al. 2009). Initial results indicate that autumn application of Plateau after fire is most effective for reducing cheatgrass (Bromus tectorum), but longer-term monitoring is needed to evaluate long-term and non-target effects. In addition to controlling invasive species, active recovery of native vegetation may be attempted. Methods may include seeding of early native colonizers such as desert globemallow, burrobrush, threeawns (Aristida spp.), and desert marigold (e.g. Abella et al. 2009, Abella et al. 2012). Increased native cover may help to reduce non-native plant invasion, helps to stabilize soils, provides a source of food and cover for wildlife, including desert tortoise (Gopherus agassizii), and provides microsites that facilitate blackbrush establishment. However, the amount of seed required for success is often prohibitive. Large-scale planting of both early colonizers and community dominants tends to be more successful in terms of plant survival, especially if outplants receive supplemental watering during the first two years (Allen 1993). Blackbrush is difficult to cultivate for outplanting due to susceptibility to fungal pathogens in the greenhouse environment.
Transition 3
State 3 to 4
This transition occurs with severe or continuous anthropogenic disturbance that increases the effects of erosion, or when the fire return interval is less than 20 years.
Additional community tables
Table 8. Community 2.1 plant community composition
| Group | Common name | Symbol | Scientific name | Annual production (lb/acre) | Foliar cover (%) | |
|---|---|---|---|---|---|---|
|
Shrub/Vine
|
||||||
| 1 | Native shrubs | 80–240 | ||||
| blackbrush | CORA | Coleogyne ramosissima | 50–120 | 8–12 | ||
| Eastern Mojave buckwheat | ERFA2 | Eriogonum fasciculatum | 3–40 | 1–3 | ||
| desert almond | PRFA | Prunus fasciculata | 0–35 | 0–2 | ||
| Acton's brittlebush | ENAC | Encelia actonii | 5–30 | 1–3 | ||
| Nevada jointfir | EPNE | Ephedra nevadensis | 1–23 | 0–2 | ||
| burrobrush | HYSA | Hymenoclea salsola | 0–20 | 0–1 | ||
| turpentine bush | ERLA12 | Ericameria laricifolia | 0–17 | 0–1 | ||
| Mojave yucca | YUSC2 | Yucca schidigera | 1–10 | 0–1 | ||
| Cooper's goldenbush | ERCO23 | Ericameria cooperi | 0–7 | 0–1 | ||
| Wiggins' cholla | CYEC3 | Cylindropuntia echinocarpa | 0–3 | 0–1 | ||
| branched pencil cholla | CYRA9 | Cylindropuntia ramosissima | 0–3 | 0–1 | ||
| narrowleaf goldenbush | ERLI6 | Ericameria linearifolia | 0–3 | 0–1 | ||
| green rabbitbrush | ERTE18 | Ericameria teretifolia | 0–1 | 0–1 | ||
| catclaw acacia | ACGR | Acacia greggii | 0–1 | 0–1 | ||
|
Tree
|
||||||
| 2 | Trees | 3–40 | ||||
| California juniper | JUCA7 | Juniperus californica | 3–40 | 1–5 | ||
|
Grass/Grasslike
|
||||||
| 3 | Native perennial grasses | 3–20 | ||||
| desert needlegrass | ACSP12 | Achnatherum speciosum | 3–20 | 0–1 | ||
| 5 | Non-native annual grasses | 0–80 | ||||
| red brome | BRRU2 | Bromus rubens | 0–80 | 0–13 | ||
| cheatgrass | BRTE | Bromus tectorum | 0–1 | 0–1 | ||
|
Forb
|
||||||
| 4 | Native forbs | 4–50 | ||||
| bristly fiddleneck | AMTE3 | Amsinckia tessellata | 0–40 | 0–4 | ||
| Forb, annual | 2FA | Forb, annual | 0–30 | 0–1 | ||
| pincushion flower | CHFR | Chaenactis fremontii | 0–20 | 0–1 | ||
| shrubby deervetch | LORI3 | Lotus rigidus | 0–15 | 0–1 | ||
| desert globemallow | SPAM2 | Sphaeralcea ambigua | 0–5 | 0–1 | ||
| wishbone-bush | MILAV | Mirabilis laevis var. villosa | 0–1 | 0–1 | ||
| chia | SACO6 | Salvia columbariae | 0–1 | 0–1 | ||
Table 9. Community 3.1 plant community composition
| Group | Common name | Symbol | Scientific name | Annual production (lb/acre) | Foliar cover (%) | |
|---|---|---|---|---|---|---|
|
Tree
|
||||||
| 1 | Trees | 12–25 | ||||
| Joshua tree | YUBR | Yucca brevifolia | 0–25 | 0–1 | ||
| California juniper | JUCA7 | Juniperus californica | 9–20 | 3–9 | ||
|
Shrub/Vine
|
||||||
| 2 | Native shrubs | 150–300 | ||||
| Eastern Mojave buckwheat | ERFA2 | Eriogonum fasciculatum | 37–113 | 3–6 | ||
| Mojave yucca | YUSC2 | Yucca schidigera | 9–66 | 2–4 | ||
| Nevada jointfir | EPNE | Ephedra nevadensis | 7–31 | 1–5 | ||
| littleleaf ratany | KRER | Krameria erecta | 2–26 | 1–2 | ||
| turpentine bush | ERLA12 | Ericameria laricifolia | 0–20 | 0–1 | ||
| burrobrush | HYSA | Hymenoclea salsola | 2–15 | 0–1 | ||
| Cooper's goldenbush | ERCO23 | Ericameria cooperi | 2–6 | 0–1 | ||
| narrowleaf goldenbush | ERLI6 | Ericameria linearifolia | 0–5 | 0–1 | ||
| Wiggins' cholla | CYEC3 | Cylindropuntia echinocarpa | 1–5 | 0–1 | ||
| Mexican bladdersage | SAME | Salazaria mexicana | 0–4 | 1–2 | ||
| turpentinebroom | THMO | Thamnosma montana | 0–4 | 1 | ||
| Acton's brittlebush | ENAC | Encelia actonii | 0–2 | 0–1 | ||
|
Grass/Grasslike
|
||||||
| 3 | Native perennial grasses | 3–20 | ||||
| desert needlegrass | ACSP12 | Achnatherum speciosum | 0–18 | 0–1 | ||
| big galleta | PLRI3 | Pleuraphis rigida | 0–12 | 0–1 | ||
| Indian ricegrass | ACHY | Achnatherum hymenoides | 0–1 | 0–1 | ||
| 5 | Non-native annual grasses | 0–20 | ||||
| red brome | BRRU2 | Bromus rubens | 0–20 | 0–8 | ||
| cheatgrass | BRTE | Bromus tectorum | 0–1 | 0–1 | ||
|
Forb
|
||||||
| 4 | Native forbs | 25–100 | ||||
| bristly fiddleneck | AMTE3 | Amsinckia tessellata | 0–80 | 0–4 | ||
| pincushion flower | CHFR | Chaenactis fremontii | 0–40 | 0–8 | ||
| shrubby deervetch | LORI3 | Lotus rigidus | 0–40 | 0–2 | ||
| chia | SACO6 | Salvia columbariae | 0–10 | 0–1 | ||
| wishbone-bush | MILAV | Mirabilis laevis var. villosa | 0–6 | 0–1 | ||
| desert globemallow | SPAM2 | Sphaeralcea ambigua | 0–5 | 0–1 | ||
| 6 | Non-native annual forbs | 0–20 | ||||
| redstem stork's bill | ERCI6 | Erodium cicutarium | 0–20 | 0–1 | ||
Interpretations
Animal community
Lizards:
Desert banded Gecko (Coleonyx variegatus variegatus)
Long-nosed leopard lizard (Gambelia wislizenii wislizenii)
Mojave collared lizard (Crotaphytus bicinctores)
Western chuckwalla (Sauromalus aster obesus)
San Diego horned lizard (Phrynosoma coronatum blainvillii)
Yellow-backed spiny lizard (Sceloporus magister uniformus)
Great Basin fence lizard (Sceloporus biseriatus longipes)
Western brush lizard (Urosaurus graciosus graciosus)
Desert side-blotched lizard (Uta stansburiana stejnegeri)
Desert night lizard (Xantusia vigilis vigilis)
Great Basin Whiptail (Aspidoscelis tigris tigris)
Snakes:
Mojave glossy snake (Arizona occidentalis candida) California kingsnake (Lampropeltis getula californae)
Red coachwhip (Masticophis flagellum piceus)
Desert night snake (Hypsiglena torquata deserticola)
California kingsnake (Lampropeltis getula californae)
Western leaf-nosed snake (Phyllorynchus decurtatus perkinsi)
Great Basin gopher snake (Pituophis catenifer deserticola)
California lyre snake (Trimorphodon biscutatus vandenburghi)
Mojave Desert sidewinder (Crotalus cerastes cerastes)
Southwestern speckled rattlesnake (Crotalus mitchelli Pyrrhus)
Red diamond rattlesnake (Crotalus ruber ruber)
The following mammals are likely to occur in this ecological site:
Long-tailed weasel (Mustela frenata latirosta)
Mammals:
Long-tailed weasel (Mustela latirosta)
California desert bat (Myotis californicus stephensi)
Western pipistrelle (Pipistrellus hesperus hesperus)
Desert big brown bat (Eptesicus fuscus pallidus)
Hoary bat (Lasiurus cinereus cinereus)
Pallid bat (Antrozous pallidus minor)
Desert coyote (Canis macrotis arsipus)
Common gray fox (Urocyon cinereoargenteus scottii)
Desert bobcat (Lynx rufus baileyi)
California ringtail (Bassariscus astutus ocatvus)
Southern mule deer (Odoceileus hemionus fuliginatus)
Desert bighorn sheep (Ovis canadensis nelson)
Southern Desert cottontail (Sylvilagus audobonii arizonae)
Desert blacktail jackrabbit (Lepus californicus deserticola)
Whitetail antelope squirrel (Ammospermphilus leucurus leucurus)
Western Mojave ground squirrel (Spermophilus beecheyi parvulus)
Pallid (San Diego) pocket mouse (Chaetodipus fallax pallidus)
Mojave little pocket mouse (Perognathus longimembris longimembris)
Long-tailed pocket mouse (Chaetodipus mojavensis)
Merriam's kangaroo rat (Dipodomys deserti)
Desert harvest mouse (Reithrodontomys megalotis megalotis)
Desert wood rat (Neotoma fuscipes simplex)
White-throated wood rat (Neotoma albigula venusta)
Desert canyon mouse (Peromyscus crinitus stephensi)
Cactus mouse (Peromyscus eremicus eremicus)
Southern brush mouse (Peromyscus boylii rowleyi)
Sonoran deer mouse (Peromyscus maniculatus sonoriensis)
Desert grasshopper mouse (Onychomys torridus pulcher)
Desert shrew (Notiosorex crawfordi crawfordi
Recreational uses
This ecological site can be used for hiking and aesthetic enjoyment. Pediment landscapes are an unusual and interesting feature of arid environments.
Wood products
California juniper is a poor source of lumber because of low volume and multi-stemmed growth form. However, early ranchers used juniper for fenceposts, and it is used for fuel and as Christmas trees (Cope 1992).
Other products
California juniper was used by Native Americans for a variety of medicinal purposes, including cold remedies, cough treatment, anticonvulsive, to induce sweating, for hangovers, for hypotension, fever and as a muscle relaxant for childbirth relief. Berries were eaten fresh, and were dried for later use, when they were ground to make porridge or to make bread. The Kawaiisu used the bark as a building cover, and used the wood to make arrows and cooking utensils. (http://herb.umd.umich.edu/herb/search.pl?searchstring=Juniperus+californica)
Blackbrush was used by the Kawaiisu for treating gonorrhea, and the Havasupai used blackbrush as source of fodder when grass was not available. (http://herb.umd.umich.edu/herb/search.pl?searchstring=Coleogyne+ramosissima).
Supporting information
Inventory data references
The following NRCS plots were used to describe this ecological site:
Community Phase 2.1:
12497-199-17(Type location)
12497-123-009
WYPT312
12497-134-01
Community Phase 3.1:
1249753025
12497-053-C
12497-053-D
12497-053-E
Type locality
| Location 1: San Bernardino County, CA | |
|---|---|
| UTM zone | N |
| UTM northing | 3770127 |
| UTM easting | 571507 |
| General legal description | The type location is approximately 3 miles from the west entrance to Joshua Tree National Park, and 1/3 mile northeast of Park Boulevard in Joshua Tree National Park. |
Other references
Abella, S. R., D. J. Craig, S. D. Smith, and A. C. Newton. 2012. Identifying native vegetation for reducing exotic species during the restoration of desert ecossytems. Restoration Ecology.
Abella, S. R. 2009. Post-fire plant recovery in the Mojave and Sonoran Deserts of western North America. Journal of Arid Environments 73:699-707.
Abella, S. R., J. L. Gunn, M. L. Daniels, J. D. Springer, and S. E. Nyoka. 2009. Using a diverse seed mix to establish native plants on a Sonoran Desert burn. Native Plants Journal 10:21-31.
Abella, S. R., E. C. Engel, C. L. Lund, and J. E. Spencer. 2009. Early post-fire establishment on a Mojave Desert burn. Madroño 56:137-148.
Abrahams, A. D. and A. Parsons. 1991. Relationship between infiltration and stone cover on a semiarid hillslope, southern Arizona. Journal of Hydrology 122:45-59.
Allen, E. B. 1993. Restoration ecology: limits and possibilities in arid and semiarid lands. Pages 7-15 in Wildland shrub and arid land restoration symposium. U.S. Department of Agriculture, Forest Service, Intermountain Research Station, Las Vegas, NV.
Beatley, J. C. 1969. Dependence of desert rodents on winter annuals and precipitation. Ecology 50:721-724.
Beatley, J. C. 1974. Effects of rainfall and temperature on the distribution and behavior of Larrea tridentata (Creosote-bush) in the Mojave Desert of Nevada. Ecology 55:245-261.
Beatley, J. C. 1976. Rainfall and fluctuating plant populations in relation to distributions and numbers of desert rodents in southern Nevada. Oecologia 24:21-42.
Belnap, J., J. H. Kaltenecker, R. Rosentreter, J. Williams, S. Leonard, and D. Eldridge. 2001. Biological soil crusts: ecology and management. Technical Reference 1730-2, United States Department of the Interior Bureau of Land Management, Denver, CO.
Bowers, J. E. 2005. Effects of drought on shrub survival and longevity in the northern Sonoran Desert. Journal of the Torrey Botanical Society 132:421-431.
Brooks, M. L. 1999. Habitat invasibility and dominance by alien annual plants in the western Mojave Desert. Biological Invasions 1:325-337.
Brooks, M. L. 2003. Effects of increased soil nitrogen on the dominance of alien annual plants in the Mojave Desert. Journal of Applied Ecology 40:344-353.
Brooks, M. L., C. M. D'Antonio, D. M. Richardson, J. B. Grace, J. E. Keeley, J. M. DiTomaso, R. J. Hobbs, M. Pellant, and D. Pyke. 2004. Effects of invasive alien plants on fire regimes. Bioscience 54:677-689.
Brooks, M. L., T. C. Esque, and T. Duck. 2007. Creosotebush, blackbrush, and interior chaparral shrublands. RMRS-GTR-202.
Brooks, M. L. and J. R. Matchett. 2006. Spatial and temporal patterns of wildfires in the Mojave Desert, 1980-2004. Journal of Arid Environments 67:148-164.
Brown, D. E. and R. A. Minnich. 1986. Fire and Changes in Creosote Bush Scrub of the Western Sonoran Desert, California. American Midland Naturalist 116:411-422.
Bull, W. B. 1997. Discontinuous ephemeral streams. Geomorphology 19:227-276.
Cope, Amy B. 1992. Juniperus californica. In: Fire Effects Information System, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer). Available: http://www.fs.fed.us/database/feis/ [2012, January 11].
D'Antonio, C. M. and P. M. Vitousek. 1992. Biological invasions by exotic grasses, the grass/fire cycle, and global change. Annual Review of Ecology and Systematics 23:63-87.
Dohrenwend, J. C. and A. J. Parsons. 2009. Pediments in arid environments. Pages 375-412 in A. J. Parsons and A. D. Abrahams, editors. Geomorphology of desert environments. Springer.
Edinger-Marshall, S. B. and L. J. Lund. 1999. Gravel dispersion on a granite pediment (East Mojave Desert, California): a short-term look at erosional processes. Earth Surface Processes and Landforms 24:349-359.
Fuhrmann, K., K. Weber, and C. Decker. 2009. Restoring burned areas at Zion National Park (Utah). Restoration Ecology 27:132-133.
Guerrero-Campo, J. and G. Montserrat-Marti. 2000. Effects of soil erosion on the floristic composition of plant communities on marl in northeast Spain. Journal of Vegetation Science 11:329-336.
Guerrero-Campo, J., S. Palacio, and G. Montserrat-Marti. 2008. Plant traits enabling survival in Mediterranean badlands in northeastern Spain suffering from soil erosion. Journal of Vegetation Science 19:457-464.
Hereford, R., R. H. Webb, and C. I. Longpre. 2006. Precipitation history and ecosystem response to multidecadal precipitation variability in the Mojave Desert region, 1893-2001. Journal of Arid Environments 67:13-34.
Matchett, J. R., A. O'Neill, M. Brooks, C. Decker, J. Vollmer, and C. Deuser. 2009. Reducing fine fuel loads, controlling invasive annual grasses, and manipulating vegetation composition in Zion Canyon, Utah. Joint Fire Science Program, El Portal, California.
Minnich, R. A. 2003. Fire and dynamics of temperature desert woodlands in Joshua Tree National Park. Contract, Joshua Tree National Park.
Miriti, M. N., S. Rodriguez-Buritica, S. J. Wright, and H. F. Howe. 2007. Episodic death across species of desert shrubs. Ecology 88:32-36.
Norton, J. B., T. A. Monaco, and U. Norton. 2007. Mediterranean annual grasses in western North America: kids in a candy store. Plant Soil 298:1-5.
Pendleton, B. K. and S. E. Meyer. 2004. Habitat-correlated variation in blackbrush (Coleogyne ramosissima: Rosaceae) seed germination response. Journal of Arid Environments 59:229-243.
Rao, L. E. and E. B. Allen. 2010. Combined effects of precipitation and nitrogen deposition on native and invasive winter annual production in California deserts. Oecologia 162:1035-1046.
Rao, L. E., E. B. Allen, and T. M. Meixner. 2010. Risk-based determination of critical nitrogen deposition loads for fire spread in southern California deserts. Ecological Applications 20:1320-1335.
Reid, C. R., S. Goodrich, and J. E. Bowns. 2006. Cheatgrass and red brome: history and biology of two invaders. Pages 27-32 in Shrublands under fire: disturbance and recovery in a changing world. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Cedar City, Utah.
Rhode, D. 2002. Early Holocene juniper woodland and chaparral taxa in the central Baja California Peninsula, Mexico. Quaternary Research 57:102-108.
Rickard, W. H. and J. C. Beatley. 1965. Canopy-coverage of the desert shrub vegetation mosaic of the Nevada test site. Ecology 46:524-529.
Sawyer, J. O., T. Keeler-Woolf, and J. M. Evans. 2009. A manual of California vegetation. 2nd edition. California Native Plant Society, Sacramento, California.
Steers, R. J. and E. B. Allen. 2010. Post-fire control of invasive plants promotes native recovery in a burned desert shrubland. Restoration Ecology 18:334-343.
Vamstad, M. S. 2009. Effects of fire on vegetation and small mammal communities in a Mojave Desert Joshua tree woodland. M.S. University of California, Riverside, Riverside, Ca.
Vasek, F. C. 1983. Plant succession in the Mojave Desert. Crossosoma 9:1-23.
Warren, S. D. and D. J. Eldridge. 2003. Biological soil crusts and livestock in arid ecosystems are they compatible? Pages 401-416 in J. Belnap and O. L. Lange, editors. Biological soil crusts: structure, function, and management. Springer-Verlag, Berlin, Germany.
Webb, R. H., M. B. Muroy, T. C. Esque, D. E. Boyer, L. A. DeFalco, D. F. Haines, D. Oldershaw, S. J. Scoles, K. A. Thomas, J. B. Blainey, and P. A. Medica. 2003. Perennial vegetation data from permanent plots on the Nevada Test Site, Nye County, Nevada. U.S. Geological Society, Tucson, AZ.
Webb, R. H., Steiger, J.W., Turner, R.M. 1987. Dynamics of Mojave Desert shrub assemblages in the Panamint Mountains, California. Ecology 68:478-490.
Contributors
Alice Miller
Allison Tokunaga
Approval
Kendra Moseley, 10/21/2024
Rangeland health reference sheet
Interpreting Indicators of Rangeland Health is a qualitative assessment protocol used to determine ecosystem condition based on benchmark characteristics described in the Reference Sheet. A suite of 17 (or more) indicators are typically considered in an assessment. The ecological site(s) representative of an assessment location must be known prior to applying the protocol and must be verified based on soils and climate. Current plant community cannot be used to identify the ecological site.
| Author(s)/participant(s) | |
|---|---|
| Contact for lead author | |
| Date | 06/15/2012 |
| Approved by | Kendra Moseley |
| Approval date | |
| Composition (Indicators 10 and 12) based on | Annual Production |
Indicators
-
Number and extent of rills:
-
Presence of water flow patterns:
-
Number and height of erosional pedestals or terracettes:
-
Bare ground from Ecological Site Description or other studies (rock, litter, lichen, moss, plant canopy are not bare ground):
-
Number of gullies and erosion associated with gullies:
-
Extent of wind scoured, blowouts and/or depositional areas:
-
Amount of litter movement (describe size and distance expected to travel):
-
Soil surface (top few mm) resistance to erosion (stability values are averages - most sites will show a range of values):
-
Soil surface structure and SOM content (include type of structure and A-horizon color and thickness):
-
Effect of community phase composition (relative proportion of different functional groups) and spatial distribution on infiltration and runoff:
-
Presence and thickness of compaction layer (usually none; describe soil profile features which may be mistaken for compaction on this site):
-
Functional/Structural Groups (list in order of descending dominance by above-ground annual-production or live foliar cover using symbols: >>, >, = to indicate much greater than, greater than, and equal to):
Dominant:
Sub-dominant:
Other:
Additional:
-
Amount of plant mortality and decadence (include which functional groups are expected to show mortality or decadence):
-
Average percent litter cover (%) and depth ( in):
-
Expected annual annual-production (this is TOTAL above-ground annual-production, not just forage annual-production):
-
Potential invasive (including noxious) species (native and non-native). List species which BOTH characterize degraded states and have the potential to become a dominant or co-dominant species on the ecological site if their future establishment and growth is not actively controlled by management interventions. Species that become dominant for only one to several years (e.g., short-term response to drought or wildfire) are not invasive plants. Note that unlike other indicators, we are describing what is NOT expected in the reference state for the ecological site:
-
Perennial plant reproductive capability:
Print Options
Sections
Font
Other
The Ecosystem Dynamics Interpretive Tool is an information system framework developed by the USDA-ARS Jornada Experimental Range, USDA Natural Resources Conservation Service, and New Mexico State University.
Click on box and path labels to scroll to the respective text.