
Natural Resources
Conservation Service
Ecological site R030XB188CA
Cool Shallow to Moderately Deep Fans
Last updated: 10/21/2024
Accessed: 04/14/2026
General information
Provisional. A provisional ecological site description has undergone quality control and quality assurance review. It contains a working state and transition model and enough information to identify the ecological site.
Figure 1. Mapped extent
Areas shown in blue indicate the maximum mapped extent of this ecological site. Other ecological sites likely occur within the highlighted areas. It is also possible for this ecological site to occur outside of highlighted areas if detailed soil survey has not been completed or recently updated.
MLRA notes
Major Land Resource Area (MLRA): 030X–Mojave Basin and Range
MLRA Description:
Major Land Resource Area (MLRA) 30, Mojave Desert, is found in southern California, southern Nevada, the extreme southwest corner of Utah and northwestern Arizona within the Basin and Range Province of the Intermontane Plateaus. The climate of the area is hot (primarily hyperthermic and thermic; however at higher elevations, generally above 5000 feet, mesic, cryic and frigid) and dry (aridic). Elevations range from below sea level to over 12,000 feet in the higher mountain areas found within the MLRA. Due to the extreme elevational range found within this MLRA, Land Resource Units (LRUs) were designated to group the MLRA into similar land units.
LRU Description:
This LRU (designated by ’XB’) is found across the eastern half of California, much of the mid-elevations of Nevada, the southernmost portions of western Utah, and the mid-elevations of northwestern Arizona. Elevations range from 1800 to 5000 feet and precipitation ranges from 4 to 9 inches per year, but is generally between 5-6 inches. This LRU is characterized primarily by the summer precipitation it receives, ranging from 18 – 35% but averages 25%. Summer precipitation falls between July and September in the form of rain, and winter precipitation falls starting in November and ends between February and March, also mostly in the form of rain; however it does receive between 0 and 3 inches of snow, with an average of 1 inch. The soil temperature regime is thermic and the soil moisture regime is typic-aridic. Vegetation includes creosote bush, burrobush, Nevada jointfir, ratany, Mojave yucca, Joshua tree, chollas, cactus, big galleta grass and several other warm season grasses. At the upper portions of the LRU, plant production and diversity are greater and blackbrush is a common dominant shrub.
Ecological site concept
This ecological site occurs on fan aprons over pediment with very shallow to shallow soils and cool-thermic soil temperatures. Slopes range from 2 to 15.
This is a group ecological concept with a provisional STM that also covers the following ecological sites: R030XB009CA, R030XB043NV, R030XB044NV, R030XB053NV, R030XB107NV, R030XB134CA, R030XB143CA, R030XB168CA, R030XB173CA, R030XB183CA, R030XE006CA, R030XE199CA, R030XE200CA, R030XG021CA
Associated sites
| R030XB170CA |
Bouldery Very Shallow To Shallow Gravelly Slopes Cool hills and mountains. The dominant species are singleleaf pinyon (Pinus monophylla), Muller oak (Quercus cornelius-mulleri) and blackbrush (Coleogyne ramosissima). |
|---|---|
| R030XB005NV |
Arid Active Alluvial Fans Fan piedmonts, alluvial fans and fan remnants. The dominant species are burrobush (Ambrosia dumosa) and creosote bush (Larrea tridentata). |
| R030XB166CA |
Dissected Pediment, Cool Cool thermic dissected pediments with very shallow soils on slopes of 4 to 30% between the elevations of 3280 to 5180 feet. Blackbrush (Coleogyne ramosissima) and California juniper (Juniperus californica) dominate. Shallow soils and high run-off make it difficult for creosote bush to establish at this site. |
| R030XB168CA |
Cool Deep Sandy Fans Cool fan aprons and fan remnants with California juniper (Juniperus californica) and blackbrush (Coleogyne ramosissima) as dominant plant species. Soils on R030XB168CA are moderately deep to very deep with very low to low runoff class. Soil temperatures may be too cool to support creosote bush (Larrea tridentata). |
| R030XB173CA |
Coarse Loamy Very Deep Fan Remnants Cool thermic fan remnants with 2 to 8% slopes between 3500 to 4600 feet in elevation. Soils are very deep sands. Blackbrush (Coleogyne ramosissima), Joshua Tree (Yucca brevifolia var. brevifolia), and big galleta (Pleuraphis rigida) dominate the site. |
| R030XB174CA |
Sandy Fan Aprons This ecological site occurs on both cool and warm thermic fan aprons at elevations of 3000 to 4600 feet. Very deep soils distinguish this site from R030XB188CA. Creosote bush (Larrea tridentata), big galleta (Pleuraphis rigida), and Joshua tree (Yucca brevifolia var. brevifolia) dominate the site. |
| R030XB183CA |
Loamy Very Deep Fan Remnants This ecological site occurs on both cool and warm thermic fan aprons and fan remnants at elevations of 3050 to 5130 feet. Deep soils with an argillic horizon distinguish this site from R030XB188CA. The site is also dominated by blackbrush (Coleoygne ramosissima)and creosote bush (Larrea tridentata). |
| R030XB192CA |
Very Rarely Flooded, Warm Thermic Fan Piedmonts Warm thermic fan aprons with very rare sheet flooding at elevations of 2100 to 4200 feet. Soils are typically very deep. The site is co-dominated by creosote bush (Larrea tridentata), desertsenna (Senna armata), and big galleta (Pleuraphis rigida). Sheet flooding supports desertsenna, which is typically associated with wash habitats. |
Similar sites
| R030XB143CA |
Shallow Granitic Loam 5-7" P.Z. This site is found on both cool thermic and warm thermic fan remnants with 2 to 30 percent slopes within 3200 to 4800 feet in elevation. Very deep soils distinguish this site from R030XB188CA. The dominant species are blackbrush (Coleogyne ramosissima), creosote bush (Larrea tridentata) and Indian ricegrass (Achnatherum hymenoides). |
|---|---|
| R030XB166CA |
Dissected Pediment, Cool Cool thermic dissected pediments with very shallow soils on slopes of 4 to 30% between the elevations of 3280 to 5180 feet. Blackbrush (Coleogyne ramosissima) and California juniper (Juniperus californica) dominate. Shallow soils and high run-off make it difficult for creosote bush (Larrea tridentata) to establish at this site. These pediments are not covered by any type of fan. |
| R030XB183CA |
Loamy Very Deep Fan Remnants This ecological site occurs on both cool and warm thermic fan aprons and fan remnants at elevations of 3050 to 5130 feet. Deep soils with an argillic horizon distinguish this site from R030XB188CA. The site is also dominated by blackbrush (Coleogyne ramosissima) and creosote bush (Larrea tridentata). |
| R030XB168CA |
Cool Deep Sandy Fans Cool fan aprons and fan remnants with California juniper (Juniperus california) and blackbrush (Coleogyne ramosissima) as dominant plant species. Soils on R030XB168CA are moderately deep to very deep with very low to low runoff class. Soil temperatures may be too cool to support creosote bush (Larrea tridentata). |
| R030XB173CA |
Coarse Loamy Very Deep Fan Remnants Cool thermic fan remnants with 2 to 8% slopes between 3500 to 4600 feet in elevation. Very deep soils distinguish this site from R030XB188CA. Blackbrush (Coleogyne ramosissima), big galleta (Pleuraphis rigida), and Joshua Tree (Yucca brevifolia var. brevifolia) dominate the site. |
| R030XB174CA |
Sandy Fan Aprons This ecological site occurs on both cool and warm thermic fan aprons at elevations of 3000 to 4600 feet. Very deep soils distinguish this site from R030XB188CA. Creosote bush (Larrea tridentata), big galleta (Pleuraphis rigida), and Joshua tree (Yucca brevifolia var. brevifolia) dominate the site. |
Table 1. Dominant plant species
| Tree |
Not specified |
|---|---|
| Shrub |
(1) Coleogyne ramosissima |
| Herbaceous |
Not specified |
Physiographic features
This ecological site occurs on pediment, fan aprons over pediment, and low hills. Elevations range from 3200 to 5130 feet, and slopes range from 2 to 15 percent. Runoff class is medium to high.
Table 2. Representative physiographic features
| Landforms |
(1)
Fan apron
(2) Hill (3) Pediment |
|---|---|
| Flooding frequency | None |
| Ponding frequency | None |
| Elevation | 3,200 – 5,130 ft |
| Slope | 2 – 15% |
| Water table depth |
Not specified |
| Aspect | Aspect is not a significant factor |
Climatic features
The climate at this site is arid characterized by cool, somewhat moist winters and hot, dry summers. The average annual precipitation ranges from 4 to 7 inches with most precipitation falling as rain from November to March. Mean annual air temperature ranges from 55 to 63 degrees Fahrenheit. June, July and August can experience average maximum temperatures of 100 degrees Fahrenheit while December and January can have average minimum temperatures near 20 degrees Fahrenheit.
Maximum and minimum monthly climate data for this ESD were generated by the Climate Summarizer
(http://www.nm.nrcs.usda.gov/technical/handbooks/nrph/Climate_Summarizer.xls) using data
from the following climate stations (results are weighted averages; numbers in square brackets represent relative weights):
LTHC1 Lost Horse, Joshua Tree National Park (Period of record = 1991 to 2011) [2]
44405 JOSHUA TREE, CA (Period of record = 1959 to 2011) [1]
44467 Kee Ranch, CA (Period of record = 1948 to 1979) [1]
45112 Yucca Valley, CA (Period of record = 1990 to 2011) [1]
The Lost Horse weather station is closest to this ecological site but is limited by the number of years data was collected. The Joshua Tree weather station is also nearby this ecological site but is located at approximately 2750 feet in elevation while the ecological site has an elevational range of 3200 to 5200 feet. This weather station is lacking precipitation data for the years between 1975 and 2008 and there is very little temperature data. Kee Ranch weather station contains precipitation data for all years of the period of record but has no temperature data. The Yucca Valley weather station contains temperature and precipitation data for the 20 year period of record.
Frost Free Period and Mean Annual Precipitation were determined using a variety of climate data and models. Freeze Free Period is a best guess based on available temperature data and the Frost Free Period. Due to sparse temperature data, the Frost Free Period determined using the Climate Summarizer does not agree with the models used to populate the National Soil Information System.
Table 3. Representative climatic features
| Frost-free period (average) | 270 days |
|---|---|
| Freeze-free period (average) | 300 days |
| Precipitation total (average) | 7 in |
Figure 2. Monthly precipitation range
Figure 3. Monthly average minimum and maximum temperature
Influencing water features
Soil features
This ecological site is found on colluvial and alluvial soils derived from gneiss, granite and granitoid rocks. Soils are very shallow and shallow over weathered, fractured bedrock. The surface textures are loamy sand or sandy loam and the subsurface textures are sandy loam, gravelly loamy sand, and coarse sandy loam. In some Desertqueen pedons there is a thin horizon (BCt from 12 to 14 inches) with an extremely gravelly loamy coarse sand texture. Rock fragments less than 3 inches in diameter compose 70 to 85 percent of the surface cover. Subsurface rock fragments (from 2 to 14 inches) greater than 3 inches in diameter range from 2 to 15 percent, with 75 percent in the BCt horizon mentioned above. Rock fragments greater than 3 inches in diameter are not typically found at this site. Weathered, extremely weakly cemented or weakly cemented, fractured bedrock extends from below the developed horizons to 59 inches.
This ecological site is associated with the following soil series: Desertqueen (loamy, mixed, superactive, thermic, shallow Typic Haplargids); and Pinecity (mixed, thermic, shallow Typic Torripsamments).
Desertqueen soils have an argillic horizon at depths of 1 to 8 inches below the surface, and have sandy loam surface textures. Pinecity soils have little horizon development, and have loamy sand surface textures. They are sandy throughout the soil profile.
This ecological site is found in the following map units of the Joshua Tree National Park Soil Survey (CA794) listed by map unit ID; map unit; component; and component percent:
3681; Morongo-Jumborox complex, 4 to 8 percent slopes, warm; Desertqueen; 4%
3684; Morongo loamy sand, 4 to 8 percent slopes, warm; Pinecity; 5%
4071; Helendale-Desertqueen association, 4 to 15 percent slopes; Desertqueen; very rarely flooded; 15%
4605; Pinecity complex, 2 to 8 percent slopes; Pinecity; 80%
4610; Jumborox-Desertqueen-Rock outcrop association, 2 to 8 percent slopes; Desertqueen; 35%; Pinecity; 5%
Table 4. Representative soil features
| Parent material |
(1)
Colluvium
–
granite
(2) Alluvium – gneiss |
|---|---|
| Surface texture |
(1) Sandy loam (2) Loamy sand |
| Family particle size |
(1) Loamy |
| Drainage class | Well drained to somewhat excessively drained |
| Permeability class | Moderately rapid to rapid |
| Soil depth | 2 – 14 in |
| Surface fragment cover <=3" | 70 – 85% |
| Surface fragment cover >3" | Not specified |
| Available water capacity (0-40in) |
0.3 – 1.4 in |
| Calcium carbonate equivalent (0-40in) |
1% |
| Electrical conductivity (0-40in) |
2 mmhos/cm |
| Sodium adsorption ratio (0-40in) |
5 |
| Soil reaction (1:1 water) (0-40in) |
6.1 – 8 |
| Subsurface fragment volume <=3" (Depth not specified) |
2 – 75% |
| Subsurface fragment volume >3" (Depth not specified) |
Not specified |
Ecological dynamics
The major factors affecting this ecological site are soil depth, soil stability, and runoff from rock outcrop. The dominant species in this ecological site are blackbrush (Coleogyne ramosissima) and creosote bush (Larrea tridentata). Both species are long-lived and are best supported in stable environments. The fractured bedrock under the shallow soils slows the downward movement of water, and holds water near the surface where blackbrush roots are concentrated. Creosote bush is typically not abundant on shallow soils. It has a wider, deeper root system with which it extracts water from deeper soils. Some roots likely penetrate the fractured bedrock for deeper water, but its abundance in this ecological site is likely assisted by runoff from adjacent rock outcrop areas. Concave surfaces often have a significant component of desertsenna (Senna armata), indicating disturbance of water through this site.
A high diversity of minor species is present. These species include Acton’s brittlebush (Encelia actonii), white ratany (Krameria grayi), California buckwheat (Eriogonum fasciculatum), Mojave yucca (Yucca schidigera), desert globemallow (Sphaeralcea ambigua), and desert needlegrass (Achnatherum speciosum). Several of these species are common on shallow or rocky soils and have a wide range of tolerance for different habitats. Desert almond (Prunus fasciculata) may also be present, particularly on concave surfaces.
The dominance of blackbrush suggests that disturbances to this area are neither common nor intense. Wildfire has historically been infrequent in the desert due to widely spaced shrubs and discontinuous fuels, but the establishment of invasive annual grasses increases wildfire risk by making a more continuous, easily ignitable fuel bed (D’Antonio and Vitousek 1992). This ecological site currently has few such species, but it is in close proximity to areas containing invasive grasses. Because invasive grasses travel easily via animal movement or recreational activities, this ecological site may be easily invaded. Invasion often begins under shrubs where nutrients and moisture allow non-native plant species establishment.
Blackbrush communities are likely to be significantly altered by fire or other widespread disturbance. The ability of blackbrush to recolonize a disturbed site is severely limited by infrequent seedling establishment (Callison and Brotherson 1985; Webb et al. 1987). Neither blackbrush (Bowns and West 1976; Hansen et al. 1999) nor creosote bush (Humphrey 1974; Brown and Minnich 1986) resprouts following a fire. Other species on this site regenerate following fire via sprouting or seed. Blackbrush is highly competitive for resources and reduced competition from late seral species for light, water, and nutrients facilitates growth of other species.
There are small areas within the ecological site where relief in the pediment surface creates microsites receiving more run-on favorable for big galleta. These areas trap fine particles creating small coppice dunes.
State and transition model
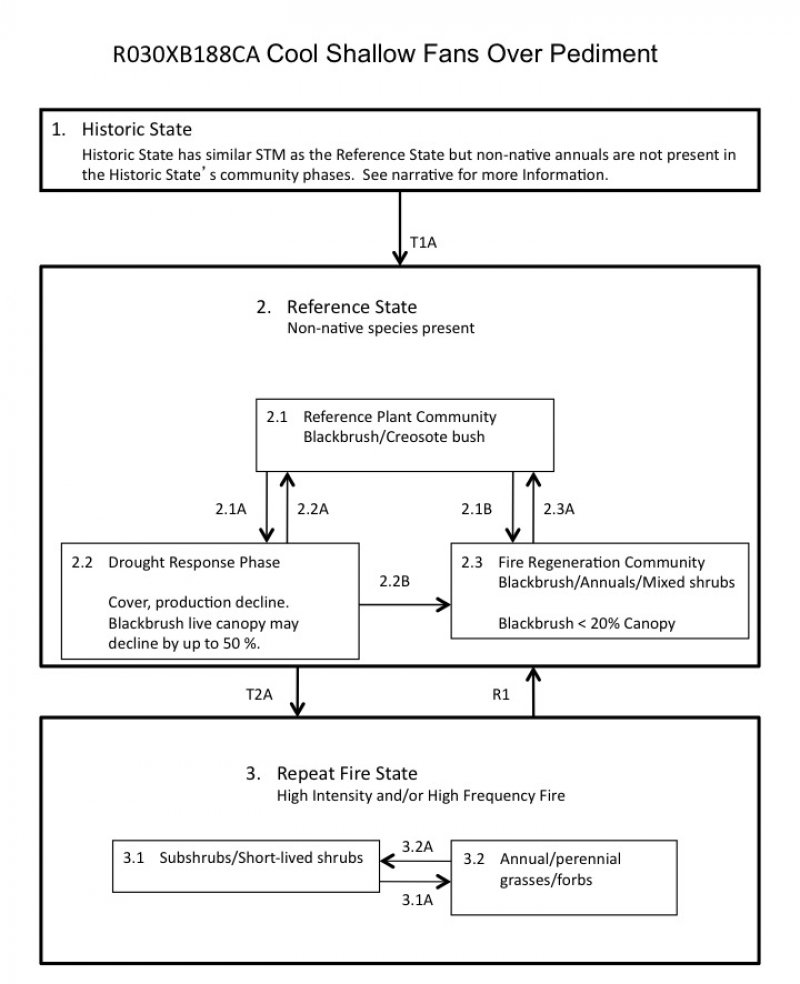
Figure 4. R030XB188CA
More interactive model formats are also available.
View Interactive Models
More interactive model formats are also available.
View Interactive Models
Click on state and transition labels to scroll to the respective text
Ecosystem states
State 2 submodel, plant communities
State 3 submodel, plant communities
State 1
Historic State
Historic-natural condition for this ecological site. This state is similar to State 2, but Historic State 1 contains only native species. If dynamics were included in this state, they would be similar to those displayed in State 2. The presence of non-native species in State 2 may increase fire frequency and intensity from that experienced in Historic State 1.
State 2
Reference State
The Reference State includes three community phases maintained by the current natural conditions for this ecological site. Indicators: Blackbrush and creosote bush are prevalent. Feedbacks: Increasing organic matter inputs, soil protected from wind erosion.
Community 2.1
Reference Plant Community

Figure 5. R030XB188CA Community Phase 2.1

Figure 6. R030XB188CA Community Phase 2.1 - no creosote bush
The reference plant community is dominated by blackbrush (Coleogyne ramosissima) and creosote bush (Larrea tridentata). Desertsenna (Senna armata) is often abundant on concave surfaces. Species diversity in this plant community is high, and includes Acton’s brittlebush (Encelia actonii), white ratany (Krameria grayi), California buckwheat (Eriogonum fasciculatum), Mojave yucca (Yucca schidigera) and desert needlegrass (Achnatherum speciosum). This ecological site is the lower elevational boundary of blackbrush and the upper elevational boundary of creosote bush, and cover and production of each species is highly variable. Several consecutive years with high precipitation could increase catclaw acacia, burrobrush, and desertsenna canopy cover. Prolonged severe drought will decrease the total canopy cover for this site (Schultz and Ostler 1995), and lead to a drought-response community phase. Years with above average precipitation produce heavy growth of native annuals and exotic grasses (Humphrey 1974; Brown and Minnich 1986), which may create a continuous fine fuel load between shrubs, which puts this community at risk for fire.
Figure 7. Annual production by plant type (representative values) or group (midpoint values)
Table 5. Annual production by plant type
| Plant type | Low (lb/acre) |
Representative value (lb/acre) |
High (lb/acre) |
|---|---|---|---|
| Shrub/Vine | 300 | 400 | 650 |
| Forb | 33 | 51 | 88 |
| Tree | 2 | 8 | 10 |
| Grass/Grasslike | 0 | 1 | 2 |
| Total | 335 | 460 | 750 |
Table 6. Ground cover
| Tree foliar cover | 0% |
|---|---|
| Shrub/vine/liana foliar cover | 25-40% |
| Grass/grasslike foliar cover | 0% |
| Forb foliar cover | 3-10% |
| Non-vascular plants | 0% |
| Biological crusts | 0% |
| Litter | 4-17% |
| Surface fragments >0.25" and <=3" | 0% |
| Surface fragments >3" | 0% |
| Bedrock | 0-4% |
| Water | 0% |
| Bare ground | 3-24% |
Community 2.2
Drought Response
This community phase is characterized by a decline in cover and production due to branch-pruning of community dominants and longer-lived minor species (including Nevada jointfir, white ratany, and Mojave yucca), and mortality of shorter-lived perennials (including burrobrush, Cooper’s goldenbush, eastern Mojave buckwheat, and big galleta), and lack of emergence of annual forbs and grasses. The dominant species of this ecological site are long-lived, drought-tolerant species that exhibit low levels of mortality during drought. Both blackbrush and creosote bush are capable of utilizing moisture at any time of the year. This ability buffers these plants from the effects of winter season drought (which is typical of this ecological site where winter precipitation is most reliable). Creosote bush germinates in response to moisture during the warm season, so may recruit if warm season rains occur during winter drought (Hereford et al. 2006). Creosote bush exhibits branch-pruning during severe drought, but mortality during drought in the Mojave Desert is very low (Webb et al. 2003, Hereford et al. 2006). Schultz and Ostler (1995) studied the effects of prolonged drought on several vegetation associations in the Northern Mojave Desert. They determined the live to dead blackbrush crown ratio after the drought, and found the low-elevation blackbrush associations had the lowest live to dead ratio of the vegetation associations they studied. The low elevation blackbrush community had 46% live crowns while the high-elevation blackbrush association had one of the highest live to dead ratios with 79% live crowns. The R030XB188CA sites are at blackbrush's lower elevational range and may experience scenarios similar to what Schultz and Ostler found at the low-elevation blackbrush association sites following prolonged severe drought. Blackbrush recruitment is episodic, and only occurs after heavy winter and spring rain (Summers et al. 2009), so recruitment will be absent during periods of drought. However, blackbrush individuals are capable of remaining dormant during periods of drought exceeding three years (Pendleton and Meyer, 2004). Blackbrush also exhibits branch-pruning during drought, with very rare mortality (Webb et al. 2003, Pendleton and Meyer, 2004).
Community 2.3
Fire regeneration community

Figure 8. R030XB188CA Community Phase 2.2
This community is characterized by decreases in blackbrush and creosote bush, and an increase in species diversity. Blackbrush survival and regeneration following fire is rare and its documentation is limited to only a few cases of blackbrush re-sprouting (Bates 1984), seedling establishment (Ellison 1950; Lei 1999), autogenic succession following fire (Thatcher 1975) and fire islands (Minnich 2003). Fires tend to remove blackbrush and creosote as dominants within these plant communities; however the low canopy cover in combination with low annual cover that can occur at these sites may only support low intensity and patchy fires. Vigorous sprouting of many other native shrubs at this site occurs following fire and the canopy cover of the respective plant species may increase (Humphrey 1953; Wallace and Romney 1972; Baldwin 1979; Callison et al. 1985; West and Hassan 1985; Conrad 1987; Minnich 1995; Loik et al. 2000; Abella et al. 2009). Islands of vegetation which were not burned by fire may provide seed sources for colonization. These unburned islands are especially important for blackbrush re-colonization. At-risk Community Phase: Community is at risk when annual plant cover creates a continuous fuel load between shrubs. Also, non-native species are limited by both water and nitrogen availability. Anthropogenic nitrogen deposition and precipitation regimes altered by climate change may accelerate the change in the Reference State’s fire ecology by further promoting the growth of non-native forbs and grasses (Dukes and Mooney 1999; Rao and Allen 2010).
Figure 9. Annual production by plant type (representative values) or group (midpoint values)
Table 7. Annual production by plant type
| Plant type | Low (lb/acre) |
Representative value (lb/acre) |
High (lb/acre) |
|---|---|---|---|
| Shrub/Vine | 150 | 175 | 200 |
| Forb | 35 | 85 | 120 |
| Tree | 1 | 2 | 3 |
| Total | 186 | 262 | 323 |
Pathway 2.1A
Community 2.1 to 2.2
This pathway occurs with prolonged or severe drought.
Pathway 2.1B
Community 2.1 to 2.3
This pathway occurs with fire.
Pathway 2.2A
Community 2.2 to 2.1
This pathway occurs with a return to average or above average precipitation.
Pathway 2.2B
Community 2.2 to 2.3
This pathway occurs with moderate to severe fire. Although live annuals are largely absent from Community Phase 2.2, standing annual biomass in drought years immediately following a period of heavy precipitation poses a severe risk for fire. Cured native annual cover may pose a risk during the first year of drought, and non-native annual grasses pose a risk for three or more years (Minnich 2003, Brooks et al. 2007, Rao et al. 2010).
Pathway 2.3a
Community 2.3 to 2.1
Community Phase 2.3 will develop into Community Phase 2.1 in the absence of fire or prolonged severe drought over time. Shrub dominance can be expected to return within 20 years but may not resemble the pre-burn composition (Callison et al. 1985).
State 3
Repeat Fire
This state develops when intense and/or frequent fire removes blackbrush and creosote bush as dominant species. There are two community phases maintained by an increase in fire intensity and/or fire frequency due to the abundance of annual species, especially invasive grasses.
Community 3.1
Subshrubs/Short-lived shrubs
This community phase develops with time without fire (5-20 years). Burrobrush and Cooper's goldenbush can quickly colonize disturbed sites from off-site seed dispersal (Tratz 1978; Web et al 1988). Burrobrush is likely to be the dominant shrub in this community phase. If the fire did not destroy underground regenerative structures, catclaw acacia (Humphrey 1953; Baldwin 1979), Mojave yucca (Loik et al. 2000;), Nevada jointfir (Wallace and Romney 1972; West and Hassan 1985), turpentinebroom (Callison et al. 1985) and water jacket (Wallace and Romney 1972; Web et al 1988) may resprout to form a stand of mixed shrubs (Conrad 1987; Minnich 1995; Abella et al. 2009). Acton's brittlebush may also colonize the area via off-site seed disperal (Conrad 1987). Others shrubs which may colonize the site following fire are snakeweed (Gutierrezia spp.) (Lei 1999) and desert globemallow (Sphaeralcea ambigua) (Abella et al. 2009). There is no data for this community phase. The concept is based on research.
Community 3.2
Annual/perennial grasses/forbs
This community phase is characterized by forbs and grasses able to colonize the site post-fire. Some of the grasses found at this site which are able to colonize an area following fire are; cheatgrass, desert needlegrass, and red brome. Forbs able to colonize the site following fire include bristly fiddleneck, pincushion flower and redstem stork's bill (Brooks and Matchett 2003; Abella et al. 2009). Canopy cover of non-native grasses and forbs, such as cheatgrass, red brome, and redstem stork's bill, may increase greater than native plants. Non-native invaders have the potential to permanently increase the fire frequency regime at these sites, maintaining this community phase (D’Antonio and Vitousek 1992; Whisenant 1990). Periodic droughts may provide native plants the opportunity to colonize the site when non-natives do not have sufficient water available to them (Brooks 2002; Bartha et al. 2003). Only cover data for this community phase is presented. There is no production data for this community phase.
Figure 10. Annual production by plant type (representative values) or group (midpoint values)
Pathway 3.1A
Community 3.1 to 3.2
This pathway occurs with fire.
Pathway 3.2a
Community 3.2 to 3.1
Overtime, community phase 3.2 will develop into community phase 3.1. Shrub dominance can be expected to return within 20 years but will not resemble the pre-burn composition (Callison et al. 1985).
Transition 1A
State 1 to 2
Non-native forbs and grasses are introduced to the Historic State. There may be only a trace amount of non-native species present. Non-native species are well adapted to the desert climate. Attempts to eradicate this species may be futile as seed sources are widespread throughout the state of California and the Southwest.
Transition T2A
State 2 to 3
High intensity fire is carried by the continuous fuel load of annual species. This transition is most likely to occur when the interstitial spaces between shrubs are filled with dried annual species, especially non-native grasses.
Restoration pathway 1
State 3 to 2
Restoration of communities severely altered by repeat fire at the landscape scale is difficult. Methods may include aerial seeding of early native colonizers such as desert globemallow, burrobrush, threeawns (Aristida spp.), and desert marigold. Increased native cover may help to reduce non-native plant invasion, helps to stabilize soils, provides a source of food and cover for wildlife, including desert tortoise (Gopherus agassizii), and provides microsites that facilitate creosote bush and blackbrush establishment. However, the amount of seed required for success is often prohibitive. Large-scale planting of both early colonizers and community dominants tends to be more successful in terms of plant survival, especially if outplants receive supplemental watering during the first two years. Creosote bush is readily cultivated for outplanting, but blackbrush is difficult to cultivate due to susceptibility to fungal pathogens in the greenhouse environment. Pre-emergent herbicides (Plateau) have been used in the year immediately post-fire to attempt to inhibit or reduce brome invasion. How successful this is on a landscape scale, and the non-target effects have not yet been determined.
Additional community tables
Table 8. Community 2.1 plant community composition
| Group | Common name | Symbol | Scientific name | Annual production (lb/acre) | Foliar cover (%) | |
|---|---|---|---|---|---|---|
|
Tree
|
||||||
| 1 | Trees | 2–10 | ||||
| Joshua tree | YUBR | Yucca brevifolia | 2–10 | 0–1 | ||
| catclaw acacia | ACGR | Acacia greggii | 0–5 | 0–1 | ||
|
Shrub/Vine
|
||||||
| 2 | Native Shrubs | 250–650 | ||||
| blackbrush | CORA | Coleogyne ramosissima | 75–350 | 8–17 | ||
| creosote bush | LATR2 | Larrea tridentata | 0–215 | 0–21 | ||
| burrobrush | HYSA | Hymenoclea salsola | 40–75 | 2–4 | ||
| white ratany | KRGR | Krameria grayi | 0–30 | 0–3 | ||
| Cooper's goldenbush | ERCO23 | Ericameria cooperi | 0–16 | 0–1 | ||
| Eastern Mojave buckwheat | ERFA2 | Eriogonum fasciculatum | 5–12 | 2–3 | ||
| Nevada jointfir | EPNE | Ephedra nevadensis | 0–12 | 0–2 | ||
| Mojave yucca | YUSC2 | Yucca schidigera | 5–10 | 0–3 | ||
| water jacket | LYAN | Lycium andersonii | 0–8 | 0–1 | ||
| Mexican bladdersage | SAME | Salazaria mexicana | 1–5 | 0–1 | ||
| Acton's brittlebush | ENAC | Encelia actonii | 0–5 | 0–1 | ||
| littleleaf ratany | KRER | Krameria erecta | 0–3 | 0–1 | ||
|
Grass/Grasslike
|
||||||
| 3 | Native Grasses | 0–2 | ||||
| big galleta | PLRI3 | Pleuraphis rigida | 0–3 | 0–1 | ||
| desert needlegrass | ACSP12 | Achnatherum speciosum | 0–1 | 0–1 | ||
|
Forb
|
||||||
| 4 | Native Forbs | 35–85 | ||||
| bristly fiddleneck | AMTE3 | Amsinckia tessellata | 0–40 | 0–5 | ||
| pincushion flower | CHFR | Chaenactis fremontii | 0–30 | 1–12 | ||
| 5 | Non-native Forbs | 0–1 | ||||
| redstem stork's bill | ERCI6 | Erodium cicutarium | 0–1 | 0–5 | ||
Table 9. Community 2.3 plant community composition
| Group | Common name | Symbol | Scientific name | Annual production (lb/acre) | Foliar cover (%) | |
|---|---|---|---|---|---|---|
|
Tree
|
||||||
| 1 | Trees | 1–3 | ||||
| blackbrush | CORA | Coleogyne ramosissima | 50–250 | – | ||
| creosote bush | LATR2 | Larrea tridentata | 50–250 | – | ||
| Eastern Mojave buckwheat | ERFA2 | Eriogonum fasciculatum | 5–40 | – | ||
| burrobrush | HYSA | Hymenoclea salsola | 5–40 | – | ||
| Mojave yucca | YUSC2 | Yucca schidigera | 10–40 | – | ||
| white ratany | KRGR | Krameria grayi | 5–30 | – | ||
| Acton's brittlebush | ENAC | Encelia actonii | 5–15 | – | ||
| littleleaf ratany | KRER | Krameria erecta | 5–15 | – | ||
| Mexican bladdersage | SAME | Salazaria mexicana | 5–15 | – | ||
| desertsenna | SEAR8 | Senna armata | 5–15 | – | ||
| Nevada jointfir | EPNE | Ephedra nevadensis | 1–10 | – | ||
| Cooper's goldenbush | ERCO23 | Ericameria cooperi | 1–10 | – | ||
| Joshua tree | YUBR | Yucca brevifolia | 1–10 | – | ||
| catclaw acacia | ACGR | Acacia greggii | 0–3 | 0–1 | ||
| California juniper | JUCA7 | Juniperus californica | 0–3 | 0–1 | ||
| turpentinebroom | THMO | Thamnosma montana | 0–2 | – | ||
| California juniper | JUCA7 | Juniperus californica | 0–1 | – | ||
| Wiggins' cholla | CYEC3 | Cylindropuntia echinocarpa | 0–1 | – | ||
| branched pencil cholla | CYRA9 | Cylindropuntia ramosissima | 0–1 | – | ||
| jojoba | SICH | Simmondsia chinensis | 0–1 | – | ||
| Mojave cottonthorn | TEST2 | Tetradymia stenolepis | 0–1 | – | ||
| catclaw acacia | ACGR | Acacia greggii | 0–1 | – | ||
| water jacket | LYAN | Lycium andersonii | 0–1 | – | ||
|
Shrub/Vine
|
||||||
| 2 | Native Shrubs | 150–200 | ||||
| creosote bush | LATR2 | Larrea tridentata | 40–45 | 4–6 | ||
| Mojave yucca | YUSC2 | Yucca schidigera | 35–45 | 2–4 | ||
| blackbrush | CORA | Coleogyne ramosissima | 5–25 | 2–12 | ||
| burrobrush | HYSA | Hymenoclea salsola | 15–25 | 0–1 | ||
| desertsenna | SEAR8 | Senna armata | 10–15 | 0–1 | ||
| Eastern Mojave buckwheat | ERFA2 | Eriogonum fasciculatum | 5–10 | 1–2 | ||
| white ratany | KRGR | Krameria grayi | 8–10 | 0–1 | ||
| turpentinebroom | THMO | Thamnosma montana | 0–5 | 0–1 | ||
| red brome | BRRU2 | Bromus rubens | 1–5 | – | ||
| Mediterranean grass | SCHIS | Schismus | 1–5 | – | ||
| Mexican bladdersage | SAME | Salazaria mexicana | 0–3 | 0–1 | ||
| jojoba | SICH | Simmondsia chinensis | 0–3 | 0–1 | ||
| Mojave cottonthorn | TEST2 | Tetradymia stenolepis | 0–3 | 0–1 | ||
| big galleta | PLRI3 | Pleuraphis rigida | 1–3 | – | ||
| Acton's brittlebush | ENAC | Encelia actonii | 0–2 | 0–1 | ||
| desert needlegrass | ACSP12 | Achnatherum speciosum | 0–2 | – | ||
| water jacket | LYAN | Lycium andersonii | 0–1 | 1–2 | ||
|
Forb
|
||||||
| 4 | Native Forbs | 30–95 | ||||
| bristly fiddleneck | AMTE3 | Amsinckia tessellata | 10–40 | 2–4 | ||
| Great Basin langloisia | LASE3 | Langloisia setosissima | 15–25 | 0–1 | ||
| pincushion flower | CHFR | Chaenactis fremontii | 5–15 | 0–1 | ||
| smooth desertdandelion | MAGL3 | Malacothrix glabrata | 0–10 | 0–3 | ||
| cryptantha | CRYPT | Cryptantha | 0–1 | 0–2 | ||
| 5 | Non-native Forbs | 5–25 | ||||
| redstem stork's bill | ERCI6 | Erodium cicutarium | 5–25 | 1–5 | ||
Table 10. Community 3.1 plant community composition
| Group | Common name | Symbol | Scientific name | Annual production (lb/acre) | Foliar cover (%) |
|---|
Table 11. Community 3.2 plant community composition
| Group | Common name | Symbol | Scientific name | Annual production (lb/acre) | Foliar cover (%) | |
|---|---|---|---|---|---|---|
|
Tree
|
||||||
| 1 | Tree | 0 | ||||
| Joshua tree | YUBR | Yucca brevifolia | 0 | 0–1 | ||
|
Shrub/Vine
|
||||||
| 2 | Native shrubs | 0 | ||||
| Mexican bladdersage | SAME | Salazaria mexicana | 0 | 0–3 | ||
| Nevada jointfir | EPNE | Ephedra nevadensis | 0 | 0–1 | ||
| burrobrush | HYSA | Hymenoclea salsola | 0 | 0–1 | ||
| peach thorn | LYCO2 | Lycium cooperi | 0 | 0–1 | ||
|
Grass/Grasslike
|
||||||
| 3 | Native Perennial Grasses | 0 | ||||
| big galleta | PLRI3 | Pleuraphis rigida | 0 | 2–8 | ||
| Indian ricegrass | ACHY | Achnatherum hymenoides | 0 | 0–1 | ||
|
Forb
|
||||||
| 4 | Native forbs | 0 | ||||
| smooth desertdandelion | MAGL3 | Malacothrix glabrata | 0 | 1–22 | ||
| small wirelettuce | STEX | Stephanomeria exigua | 0 | 0–7 | ||
| pincushion flower | CHFR | Chaenactis fremontii | 0 | 0–4 | ||
| Forb, annual | 2FA | Forb, annual | 0 | 0–2 | ||
| bristly fiddleneck | AMTE3 | Amsinckia tessellata | 0 | 0–2 | ||
| whitemargin sandmat | CHAL11 | Chamaesyce albomarginata | 0 | 0–1 | ||
| Great Basin langloisia | LASE3 | Langloisia setosissima | 0 | 0–1 | ||
Interpretations
Animal community
Both blackbrush and creosote bush are poor forage for livestock and most wildlife (Humphrey 1950, Humphrey 1953, Sampson and Jespersen 1963, Kufeld et. al 1973, Timmermann 1977, Bates 1983, Downum et. al 1989). Blackbrush provides poor to good forage for domestic goats (Sampson and Jespersen 1963, Provenza et. al 1983). Small mammals and birds eat blackbrush seeds (Mozingo 1987, West 1983).
Bighorn sheep and mule deer may tend to use this site in the winter to feed on blackbrush, desert needlegrass and big galleta (Bradley 1965, Stark 1966, Bowns and West 1976, West 1983, Mozingo 1987, Loope et. al 1988, Urness and Austin 1989, Seegmiller et. al 1990, Krausman et. al 1997). Black-tailed jackrabbits and desert woodrats will also browse creosote bush (Meyer 1974, Hoagland 1992). Desert reptiles and amphibians, such as the desert tortoise and the red-spotted toad, use creosote as a food source and shelter (Baxter 1988, Christensen 1992) but it is unlikely that desert tortoise will make use of this shallow ecological site. Small mammals such as the banner-tailed and Merriam’s kangaroo rats also use creosote bush for cover (Monson and Kessler 1940). A list of the many animals which may use the area, either temporarily or permanently, is presented below.
MAMMALS:
Badgers, Skunks, Weasels
Long-tailed Weasel (Mustela frenata latirosta)
Bats
California Desert Bat (Myotis californicus stephensi)
Western Pipistrelle (Pipistrellus hesperus hesperus)
Desert Big Brown Bat (Eptesicus fuscus pallidus)
Pallid Bat (Antrozous pallidus minor)
Bears
California Black Bear (Ursus Americanus californianus)
Canids
Desert Coyote (Canis latrans mearnsi)
Deer & Sheep
Southern Mule Deer [Blacktail] (Odocoileus hemionus fuliginatus)
Desert Bighorn Sheep (Ovis canadensis nelsoni)
Rabbits & Hares
Southern Desert Cottontail (Sylvilagus audubonii arizonae)
Rodents
Whitetail Antelope Squirrel (Ammospermophilus leucurus leucurus)
Long-tailed Pocket Mouse (Chaetodipus formosus mojavensis)
Merriam’s Kangaroo Rat (Dipodomys merriami merriami)
Desert Wood Rat (Neotoma lepida lepida)
Eastern Dusky-footed Wood Rat (Neotoma fuscipes simplex)
Sonoran Deer Mouse (Peromyscus maniculatus sonoriensis)
Desert Grasshopper Mouse (Onychomys torridus pulcher)
Shrews
Desert Shrew [Gray] (Notiosorex crawfordi crawfordi)
REPTILES:
Lizards
Yellow-backed Spiny Lizard (Sceloporus magister uniformis)
Great Basin Fence Lizard (Sceloporus biseriatus longipes)
Western Brush Lizard (Urosaurus graciosus graciosus)
Desert Side-blotched Lizard (Uta stansburiana stejnegeri)
Desert Night Lizard (Xantusia vigilis vigilis)
Western Red-tailed Skink (Eumeces gilberti rubricaudatus)
San Diego Alligator Lizard (Elgaria multicarinata webbii)
Silvery Legless Lizard (Anniella pulchra pulchra)
Snakes
California Kingsnake (Lampropeltis getula californiae)
Great Basin Gopher Snake (Pituophis catenifer deserticola)
Western Long-nosed Snake (Rhinocheilus lecontei lecontei)
Smith’s Black-headed Snake (Tantilla hobartsmithi)
Red Diamond Rattlesnake (Crotalus ruber ruber)
This list is not intended to be an exhaustive list of animals found in this ecological site. Many birds are likely to use this ecological site. This list is based on known habitat preferences (National Park Service 2012).
Recreational uses
This ecological site may be used for hiking and aesthetic enjoyment.
Other products
The Cahuilla use creosote stems and leaves to make a medicinal tea. A solution may be applied to open wounds to draw out poisons (http://www.malkimuseum.org/garden.htm).
The Kawaiisu used a decoction of blackbrush bark for treating gonorrhea Drug (http://herb.umd.umich.edu/herb/search.pl?searchstring=Coleogyne+ramosissima.
The Havasupai used blackbrush as source of fodder when grass was not available (http://herb.umd.umich.edu/herb/search.pl?searchstring=Coleogyne+ramosissima).
Supporting information
Inventory data references
Cover data for this ecological site was described using line-point intercept transects. The complete protocol for this sampling method is found in Monitoring Manual for Grassland, Shrubland and Savanna Ecosystems, Volume 1: Quick Start.
Below are the User Pedon ID locations that were used to describe each community phase.
Community Phase 2.1
12497-125-C; 33116H109; 33116H110
Type locality
| Location 1: San Bernardino County, CA | |
|---|---|
| Township/Range/Section | T2S R9E SN/A |
| UTM zone | N |
| UTM northing | 3761715 |
| UTM easting | 587694 |
| General legal description | The type location is located approximately 0.45 miles from the most easterly loop of Jumbo Rocks Campground in Joshua Tree National Park. |
Other references
Abella, S.R., E.C. Engel, C.L. Lund and J.E. Spencer. 2009. Early post-fire plant establishment on a Mojave Desert burn. Madroño 56: 137-148.
Baldwin, R.F. 1979. The effects of fire upon vegetation in Joshua Tree National Monument. Senior thesis report. Santa Barbara, CA: University of California. Unpublished paper on file at: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory, Missoula, MT. 76 p.
Bartha, S., S.J. Meiners, S.T.A. Pickett, and M.L. Cadenasso. 2003. Plant colonization
windows in a mesic old field succession. Applied Vegetation Science 6:205–212.
Bates, P.A. 1983. Prescribed burning blackbrush for deer habitat improvement. Cal-Neva Wildlife Transactions. [Volume unknown]: 174-182.
Bates, P.A. 1984. The role and use of fire in blackbrush (Coleogyne ramosissima Torr.)
communities in California. Doctoral dissertation, University of California, Davis. 56 pp.
Baxter, R.J. 1988. Spatial distribution of desert tortoises (Gopherus agassizii) at Twentynine Palms, California: implications for relocations. In: Szaro, R.C., K.E. Severson and D.R. Patton, technical coordinators. Management of amphibians, reptiles, and small mammals in North America: Proceedings of the symposium; 1988 July 19-21; Flagstaff, AZ. Gen. Tech. Rep. RM-166. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: 180-189.
Bowns, J.E. and N.E. West. 1976. Blackbrush (Coleogyne ramosissima Torr.) on southwestern Utah rangelands. Research Report 27. Logan, UT: Utah State University, Utah Agricultural Experiment Station. 27 p.
Bradley, W.G. 1965. A study of the blackbrush plant community of the Desert Game Range. Transactions, Desert Bighorn Council. 11: 56-61.
Brooks, M.L. 2002. Peak fire temperatures and effects on annual plants in the Mojave Desert. Ecological Applications 12:1088–1102.
Brooks, M.L. and J.R. Matchett. 2003. Plant community patterns in unburned and burned blackbrush (Coleogyne ramosissima Torr.) shrublands in the Mojave Desert. West. N. Am. Nat. 63, 283-298.
Brooks, M. L., T. C. Esque, and T. Duck. 2007. Creosotebush, blackbrush, and interior chaparral shrublands. RMRS-GTR-202.
Brown, D.E. and R.A. Minnich. 1986. Fire and changes in creosote bush scrub of the western Sonoran Desert, California. American Midland Naturalist 116:411–422.
Callison, J. and J.D. Brotherson. 1985. Habitat relationships of the blackbrush community (Coleogyne ramosissima) of southwestern Utah. The Great Basin Naturalist. 45(2): 321-326.
Callison, J., J.D. Brotherson and J.E. Bowns. 1985. The effects of fire on the blackbrush [Coleogyne ramosissima] community of southwestern Utah. Journal of Range Management. 38(6): 535-538.
Christensen, J. 1992. Sin City's luck tortoise. Nature Conservancy. 42(4): 8-13.
Conrad, C.E. 1987. Common shrubs of chaparral and associated ecosystems of southern California. Gen. Tech. Rep. PSW-99. Berkeley, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station. 86 p.
D’Antonio, C.M. and P.M. Vitousek. 1992. Biological invasions by exotic grasses, the grass/fire cycle, and global change. Annual Review of Ecology and Systematics 3:63–87.
Downum, K.R., S. Villegas, E. Rodriguez and D.J. Keil. 1989. Plant photosensitizers: a survey of their occurrence in arid and semiarid plants from North America. Journal of Chemical Ecology. 15(1): 345-355.
Dukes J.S. and H.A. Mooney. 1999. Does global change increase the success of biological invaders? Trends Ecol Evol 14:135–139
Ellison, L. 1950. Blackbrush burning. Unpublished report, US Forest Service, Intermountain Forest and Range Experiment Station, Ogden, UT. 5 pp.
Hansen, D.J., W.K. Ostler and D.B. Hall. 1999. The transition from Mojave Desert to Great Basin Desert on the Nevada Test Site. In: McArthur, E. Durant; Ostler, W. Kent; Wambolt, Carl L., compilers. Proceedings: shrub ecotones; 1998 August 12-14; Ephraim, UT. Proceedings RMRS-P-11. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: 148-158.
Hereford, R., R. H. Webb, and C. I. Longpre. 2006. Precipitation history and ecosystem response to multidecadal precipitation variability in the Mojave Desert region, 1893-2001. Journal of Arid Environments 67:13-34.
Hoagland, D.B. 1992. Feeding ecology of an insular population of the black-tailed jackrabbit (Lepus californicus) in the Gulf of California. The Southwestern Naturalist. 37(3): 280-286.
Humphrey, R.R. 1950. Arizona range resources. II. Yavapai County. Bull. 229. Tucson, AZ: University of Arizona, Agricultural Experiment Station. 55 p.
Humphrey, R.R. 1953. Forage production on Arizona ranges: III. Mohave County: A study in range condition. Bulletin 244. Tucson, AZ: University of Arizona, Agricultural Experiment Station. 79 p.
Humphrey, R.R. 1974. Fire in the deserts and desert grassland of North America. In: Kozlowski, T.T. and C.E. Ahlgren, eds. Fire and ecosystems. New York: Academic Press: 365-400.
Krausman, P.R., A.J. Kuenzi, R.C. Etchberger, [and others]. 1997. Diets of mule deer. Journal of Range Management. 50(5): 513-522.
Kufeld, R.C., O.C. Wallmo and C. Feddema. 1973. Foods of the Rocky Mountain mule deer. Res. Pap. RM-111. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. 31 p.
Lei, S.A. 1999. Postfire woody vegetation recovery and soil properties in blackbrush (Coleogyne ramosissima) shrubland ecotones. Journal of the Arizona-Nevada Academy of Science 32:105–115.
Loik, M.E., C.D St. Onge and J. Rogers. 2000. Post-fire recruitment of Yucca brevifolia and Yucca schidigera in Joshua Tree National Park, California. In: Keeley, Jon E.; Baer-Keeley, Melanie; Fotheringham, C. J., eds. 2nd interface between ecology and land development in California. U.S. Geological Survey: Open-File Report 00-62. Sacramento, CA: U.S. Department of the Interior, Geological Survey, Western Ecological Research Center: 79-85.
Loope, L.L., P.G. Sanchez, P.W. Tarr, [and others]. 1988. Biological invasions of arid land nature reserves. Biological Conservation. 44: 95-118.
Lovich, J.E. and D. Bainbridge. 1999. Anthropogenic degradation of the southern California desert ecosystem and prospects for natural recovery and restoration. Environmental Management 24:309–326.
Meyer, E.R. 1974. A reconnaissance survey of pollen rain in Big Bend National Park, Texas: modern control for a paleoenvironmental study. In: Wauer, R.H. and D.H. Riskind, eds. Transactions of the symposium on the biological resources of the Chihuahuan Desert region, United States and Mexico; 1974 October 17-18; Alpine, TX. Transactions and Proceedings Series No. 3. Washington, DC: U.S. Department of the Interior, National Park Service: 115-123.
McLaughlin, S.P. and J.E. Bowers. 1982. Effects
of wildfire on a Sonoran Desert plant community. Ecology 63:246–248.
Minnich, R.A. 1995. Wildland fire and early postfire succession in Joshua tree woodland and blackbrush scrub of the Mojave Desert of California. San Bernardino County Museum Association Quarterly 42:99–106.
Minnich, R.A. 2003. Fire and dynamics of temperate desert woodlands in Joshua Tree National Park. Report submitted to the National Park Service, Joshua Tree National Park. Contract number P8337000034/0001. 32 p.
Monson, G. and W. Kessler, 1940. Life history notes on the banner-tailed kangaroo rat, Merriam's kangaroo rat, and white-throated wood rat in Arizona and New Mexico. Journal of Wildlife Management. 4(1): 37-43.
Mozingo, H.N. 1987. Shrubs of the Great Basin: A natural history. Reno, NV: University of Nevada Press. 342 p.
Pendleton, B. K. and S. E. Meyer. 2004. Habitat-correlated variation in blackbrush (Coleogyne ramosissima: Rosaceae) seed germination response. Journal of Arid Environments 59:229-243.
Provenza, F.D., J.E. Bowns, P.J. Urness, [and others]. 1983. Biological manipulation of blackbrush by goat browsing. Journal of Range Management. 36(4): 513-518.
Rao, L.E. and E.B. Allen. 2010. Combined effects of precipitation and nitrogen deposition on native and invasive winter annual production in California deserts. Oecologia 162:1035–1046.
Rao, L. E., E. B. Allen, and T. M. Meixner. 2010. Risk-based determination of critical nitrogen deposition loads for fire spread in southern California deserts. Ecological Applications 20:1320-1335.
Sampson, A.W. and B.S. Jespersen. 1963. California range brushlands and browse plants. Berkeley, CA: University of California, Division of Agricultural Sciences, California Agricultural Experiment Station, Extension Service. 162 p.
Schultz, B.W. and W.K. Ostler. 1995a. Effects of prolonged drought on vegetation associations in the northern Mojave Desert. In: Roundy, B.A., McArthur, E.D., Haley, J.S., Mann, D.K. (Compilers), Proceedings: Wildland Shrub and Arid Land Restoration Symposium. pp. 228–235. Las Vegas, Nevada: October 19–21, 1993. US Department of Agriculture and Forest Servey, INT R 315.
Seegmiller, R.F., P.R. Krausman, W.H. Brown and F.M. Whiting. 1990. Nutritional composition of desert bighorn sheep forage in the Harquahala Mountains, Arizona. Desert Plants. 10(2): 87-90.
Stark, N. 1966. Review of highway planting information appropriate to Nevada. Bull. No. B-7. Reno, NV: University of Nevada, College of Agriculture, Desert Research Institute. 209 p. In cooperation with: Nevada State Highway Department.
Summers, H. A., B. N. Smith, and L. D. Hansen. 2009. Comparison of respiratory and growth characteristics of two co-occurring shrubs from a cold desert, Coleogyne ramosissima (blackbrush) and Atriplex confertifolia (shadscale). Journal of Arid Environments 73:1-6.
Thatcher, A.P. 1975. The amount of blackbrush in the natural plant community is largely controlled by edaphic conditions. Pages 155–156 in H.C. Stutz, editor, Proceedings wildland shrubs: symposium and workshop, US Forest Service, Shrub Sciences Laboratory, Provo, UT.
Timmermann, B.N. 1977. Practical uses of Larrea. In: Mabry, T.J., J.H. Hunziker and D.R. DiFeo, Jr., eds. Creosote bush: Biology and chemistry of Larrea in New World deserts. U.S./IBP Synthesis Series 6. Stroudsburg, PA: Dowden, Hutchinson & Ross, Inc: 252-256.
Tratz, W.M. 1978. Postfire vegetational recovery, productivity, and herbivore utilization of a chaparral-desert ecotone. Los Angeles, CA: California State University. 133 p. Thesis.
Urness, P.J. and D.D. Austin. 1989. The effects of grazing and browsing animals on wildlife habitats. Utah Science. 50(2): 104-107.
Wallace, A. and E.M. Romney. 1972. Radioecology and ecophysiology of desert plants at the Nevada Test Site. Rep. TID-25954. [Washington, DC]: U.S. Atomic Energy Commission, Office of Information Services. 439 p.
Webb, R.H., J.W. Steiger and R.M. Turner. 1987. Dynamics of Mojave Desert shrub assemblages in the Panamint Mountains, California. Ecology. 68(3): 478-490; 1987.
Webb, R.H., J.W. Steiger and E.B. Newman. 1988. The response of vegetation to disturbance in Death Valley National Monument, California. U.S. Geological Survey Bulletin 1793. Washington, DC: U.S. Department of the Interior, U.S. Geological Survey. 69 p.
Webb, H.W., T.C. Esque, P.A. Medica, L.A. DeFalco and M.B. Murov. 2001. Monitoring of Ecosystem Dynamics in the Mojave Desert: the Beatley Permanent Plots. US Department of the Interior, Geological Survey. FS-040-01.
Webb, R. H., M. B. Muroy, T. C. Esque, D. E. Boyer, L. A. DeFalco, D. F. Haines, D. Oldershaw, S. J. Scoles, K. A. Thomas, J. B. Blainey, and P. A. Medica. 2003. Perennial vegetation data from permanent plots on the Nevada Test Site, Nye County, Nevada. U.S. Geological Society, Tucson, AZ.
West, N.E. 1983. Colorado plateau-Mohavian blackbrush semi-desert. In: West, Neil E., ed. Temperate deserts and semi-deserts. New York: Elsevier Scientific Publishing Company: 399-411. (Goodall, David W., ed. in chief; Ecosystems of the world; vol. 5).
West, N.E. and M.A. Hassan. 1985. Recovery of sagebrush-grass vegetation following wildfire. Journal of Range Management. 38(2): 131-134.
Whisenant S.G. 1990. Changing Fire Frequencies on Idaho’s Snake River Plains: Ecological and Management Implications. Logan (UT): US Department of Agriculture, Forest Service, Intermountain Research Center. General Technical Report INT-276.
Contributors
Allison Tokunaga
Dustin Detweiler
Approval
Kendra Moseley, 10/21/2024
Rangeland health reference sheet
Interpreting Indicators of Rangeland Health is a qualitative assessment protocol used to determine ecosystem condition based on benchmark characteristics described in the Reference Sheet. A suite of 17 (or more) indicators are typically considered in an assessment. The ecological site(s) representative of an assessment location must be known prior to applying the protocol and must be verified based on soils and climate. Current plant community cannot be used to identify the ecological site.
| Author(s)/participant(s) | |
|---|---|
| Contact for lead author | |
| Date | 10/22/2024 |
| Approved by | Kendra Moseley |
| Approval date | |
| Composition (Indicators 10 and 12) based on | Annual Production |
Indicators
-
Number and extent of rills:
-
Presence of water flow patterns:
-
Number and height of erosional pedestals or terracettes:
-
Bare ground from Ecological Site Description or other studies (rock, litter, lichen, moss, plant canopy are not bare ground):
-
Number of gullies and erosion associated with gullies:
-
Extent of wind scoured, blowouts and/or depositional areas:
-
Amount of litter movement (describe size and distance expected to travel):
-
Soil surface (top few mm) resistance to erosion (stability values are averages - most sites will show a range of values):
-
Soil surface structure and SOM content (include type of structure and A-horizon color and thickness):
-
Effect of community phase composition (relative proportion of different functional groups) and spatial distribution on infiltration and runoff:
-
Presence and thickness of compaction layer (usually none; describe soil profile features which may be mistaken for compaction on this site):
-
Functional/Structural Groups (list in order of descending dominance by above-ground annual-production or live foliar cover using symbols: >>, >, = to indicate much greater than, greater than, and equal to):
Dominant:
Sub-dominant:
Other:
Additional:
-
Amount of plant mortality and decadence (include which functional groups are expected to show mortality or decadence):
-
Average percent litter cover (%) and depth ( in):
-
Expected annual annual-production (this is TOTAL above-ground annual-production, not just forage annual-production):
-
Potential invasive (including noxious) species (native and non-native). List species which BOTH characterize degraded states and have the potential to become a dominant or co-dominant species on the ecological site if their future establishment and growth is not actively controlled by management interventions. Species that become dominant for only one to several years (e.g., short-term response to drought or wildfire) are not invasive plants. Note that unlike other indicators, we are describing what is NOT expected in the reference state for the ecological site:
-
Perennial plant reproductive capability:
Print Options
Sections
Font
Other
The Ecosystem Dynamics Interpretive Tool is an information system framework developed by the USDA-ARS Jornada Experimental Range, USDA Natural Resources Conservation Service, and New Mexico State University.
Click on box and path labels to scroll to the respective text.