
Natural Resources
Conservation Service
Ecological site R030XE200CA
Xeric Very Deep Sandy Fan Aprons On Pediments
Last updated: 10/21/2024
Accessed: 12/22/2024
General information
Provisional. A provisional ecological site description has undergone quality control and quality assurance review. It contains a working state and transition model and enough information to identify the ecological site.
Figure 1. Mapped extent
Areas shown in blue indicate the maximum mapped extent of this ecological site. Other ecological sites likely occur within the highlighted areas. It is also possible for this ecological site to occur outside of highlighted areas if detailed soil survey has not been completed or recently updated.
MLRA notes
Major Land Resource Area (MLRA): 030X–Mojave Basin and Range
MLRA Description:
Major Land Resource Area (MLRA) 30, Mojave Desert, is found in southern California, southern Nevada, the extreme southwest corner of Utah and northwestern Arizona within the Basin and Range Province of the Intermontane Plateaus. The climate of the area is hot (primarily hyperthermic and thermic; however at higher elevations, generally above 5000 feet, mesic, cryic and frigid) and dry (aridic). Elevations range from below sea level to over 12,000 feet in the higher mountain areas found within the MLRA. Due to the extreme elevational range found within this MLRA, Land Resource Units (LRUs) were designated to group the MLRA into similar land units.
LRU Description:
This LRU (designated by ’XE’) is found only in California at the transition zone between MLRA 20, the Southern California Mountains, and MLRA 30. Elevations range from 3500 to 5800 feet and precipitation ranges from 8 to 12 inches per year. The LRU is characterized primarily by cool thermic and mesic soil temperature regimes and aridic bordering on xeric soil moisture regimes. Precipitation is mostly winter, receiving approximately 85% between October and February, going from rain in the fall to snow into the winter. Snow can range from between 1 and 6 inches. Soils show indications of greater moisture storage with some mollic colors in the soil profile. Vegetation is highly productive and vigorous for the Mojave Desert and includes non-typical Mojave species such as, scrub oaks, manzanita, sumac, ziziphus, mountain mahogany, presence of singleleaf pinyon, and desert needlegrass.
Classification relationships
Pinus monophylla Woodland Alliance (Sawyer et al. 2009).
Ecological site concept
This site is associated with group concept R030XB188CA.
This site occurs on undulating fan aprons on pediments at elevations of approximately 4400 to 5200 feet and slopes of 4 to 8 percent. Soils are typically very deep sands, but exposed bedrock and rock outcrops are common.
Single-leaf pinyon pine (Pinus monophylla), Muller’s oak (Quercus cornelius-mulleri), and California juniper (Juniperus californica) dominate the overstory, and blackbrush (Coleogyne ramosissima) is the dominant low shrub. Production Reference Value (RV) is 422 pounds per acre. Deep sandy soils, increased runon from exposed bedrock and outcrops, and the xeric soil moisture regime increases the competitive ability of single-leaf pinyon pine, Muller’s oak, and California juniper relative to blackbrush, and increases site suitability for other shrub species. Thus, this site has relatively high species evenness and shrub diversity.
Data ranges in the physiographic data, climate data, water features, and soil data sections of this Ecological Site Description are based on major components only (15 percent of map unit or greater).
Associated sites
| R030XE196CA |
Sandy Xeric-Intergrade Slopes R030XE196CA occurs on adjacent hill and mountain slopes. Single-leaf pinyon pine (Pinus monophylla) and Muller's oak (Quercus cornelius-mulleri) are dominant over a diverse shrub understory. |
|---|---|
| R030XY202CA |
Very Rarely To Rarely Flooded Thermic Ephemeral Stream R030XY202CA occurs on adjacent, small, rarely flooded ephemeral drainageways. California jointfir (Ephedra californica) adn burrobrush (Hymenoclea salsola) are dominant species of a diverse, productive shrub community. |
Similar sites
| R030XB189CA |
Shallow Cool Hills R030XB189CA occurs on steeper hill and mountain slopes with shallow soils. Blackbrush (Coleogyne ramosissima) and California juniper (Juniperus californica) are dominant species. Single-leaf pinyon pine (Pinus monophylla) and Muller's oak (Quercus cornelius mulleri) are trace if present. |
|---|---|
| R030XE196CA |
Sandy Xeric-Intergrade Slopes R030XE196CA occurs on steep mountain slopes with shallow soils. Blackbrush (Coleogyne ramosissima) is a minor species if present. |
| R030XE191CA |
Dry Sandy Mountain Slopes R030XE191CA occurs on steep mountain slopes with shallow soils. Blackbrush (Coleogyne ramosissima) and California juniper (Juniperus californica) are not important species. |
| R030XB168CA |
Cool Deep Sandy Fans R030XB168CA occurs on fan aprons and fan remnants with a typic aridic soil moisture regime. Production is higher, and blackbrush (Coleogyne ramosissima) and California juniper (Juniperus californica) dominate. |
| R030XB166CA |
Dissected Pediment, Cool R030XB166CA occurs on pediment with very shallow to shallow soils and a typic aridic soil moisture regime. Production is lower, and blackbrush (Coleogyne ramosissima) and California juniper (Juniperus californica) are dominant species. |
Table 1. Dominant plant species
| Tree |
(1) Pinus monophylla |
|---|---|
| Shrub |
(1) Coleogyne ramosissima |
| Herbaceous |
Not specified |
Physiographic features
This ecological site occurs on fan aprons on pediments and pediments at elevations of 4490 to 5130 feet with slopes of 4 to 8 percent. The site experiences no flooding or ponding, and runoff class is very low.
Table 2. Representative physiographic features
| Landforms |
(1)
Fan apron
(2) Pediment |
|---|---|
| Flooding frequency | None |
| Ponding frequency | None |
| Elevation | 1,369 – 1,564 m |
| Slope | 4 – 8% |
| Aspect | Aspect is not a significant factor |
Climatic features
The climate at this site is arid characterized by cool, somewhat moist winters and hot, dry summers. The average annual precipitation ranges from 7 to 10 inches with most falling as rain from November to March. Mean annual air temperature ranges from 55 to 63 degrees F, and the frost freeze period is 210 to 270 days. June, July, and August can experience average maximum temperatures of 100 degrees F while December and January can have average minimum temperatures near 20 degrees F.
Maximum and minimum monthly climate data for this ESD were generated by the Climate Summarizer
(http://www.nm.nrcs.usda.gov/technical/handbooks/nrph/Climate_Summarizer.xls) using data
from the following climate stations (results are weighted averages; numbers in square brackets represent relative weights):
LTHC1 Lost Horse, Joshua Tree National Park
(Period of record = 1991 to 2011) [1]
44467 Kee Ranch, CA (Period of record = 1948 to 1979) [1]
45112 Yucca Valley, CA (Period of record = 1990 to 2011) [2]
45863 Morongo Valley, CA (Period of record = 1948 to 1972) [2]
The combination of these stations were used because of a lack of data. The Lost Horse weather station is closest to this ecological site but is limited by the number of years data was collected. Kee Ranch weather station contains precipitation data for all years of the period of record but has no temperature data. The Yucca Valley weather station contains reliable temperature and precipitation data for the 20 year period of record at an elevation of 3260 ft. Morongo Valley, at elevation 2570 ft, only has precipitation data. This ecological site is generally found at elevations higher than these climate stations.
Table 3. Representative climatic features
| Frost-free period (average) | 270 days |
|---|---|
| Freeze-free period (average) | |
| Precipitation total (average) | 254 mm |
Figure 2. Monthly precipitation range
Figure 3. Monthly average minimum and maximum temperature
Influencing water features
Soil features
The soils associated with this ecological site formed in alluvium derived from granitoid and/or alluvium derived from gneiss over residuum weathered from granitoid and/or residuum weathered from gneiss. Soils are typically very deep over weathered bedrock. Surface textures are sand, with sand in the subsurface. Surface rock fragments less than or equal to three inches in diameter are typically about 10 percent, with no larger fragments, Subsurface fragments less than or equal to three inches in diameter are about 2 percent (for a depth of 0 to 79 inches), with no larger fragments.
The associated soil that is greater than 15 percent of any one map unit is: Thunderclap (mixed, thermic Xeric Torripsamments). This site is associated with minor components of Smithcanyon (mixed, thermic, shallow Xeric Torripsamments); and Stubbespring (loamy, mixed, superactive, thermic, shallow Xeric Haplargids) soils. The Smithcanyon and Stubbespring soils typically occur on backslopes of hills and mountains and are very shallow to shallow; where this ecological site is associated with them, they occur on low hills. The Stubbespring soils have sandy loam surface textures, and have an argillic horizon, which is not typical for this ecological site.
This ecological site is correlated with the following map units and soil components in the Joshua Tree National Park Soil Survey:
4630;Thunderclap-Smithcanyon complex, 4 to 15 percent slopes;Thunderclap;;50; Smithcanyon; moderately steep;8; Stubbespring;;5
3293;Smithcanyon-Pinecity association, 15 to 50 percent slopes;Thunderclap;rarely flooded;3
Table 4. Representative soil features
| Parent material |
(1)
Alluvium
–
granite
(2) Residuum – gneiss |
|---|---|
| Surface texture |
(1) Sand |
| Family particle size |
(1) Sandy |
| Drainage class | Somewhat excessively drained |
| Permeability class | Rapid |
| Soil depth | 152 – 203 cm |
| Surface fragment cover <=3" | 10% |
| Surface fragment cover >3" | 0% |
| Available water capacity (0-101.6cm) |
5.08 – 7.87 cm |
| Calcium carbonate equivalent (0-101.6cm) |
0 – 1% |
| Electrical conductivity (0-101.6cm) |
0 – 2 mmhos/cm |
| Sodium adsorption ratio (0-101.6cm) |
0 – 4 |
| Soil reaction (1:1 water) (0-101.6cm) |
7.4 – 8.4 |
| Subsurface fragment volume <=3" (Depth not specified) |
2% |
| Subsurface fragment volume >3" (Depth not specified) |
0% |
Ecological dynamics
This site occurs on undulating fan aprons over pediments, typically with very deep soils, at elevations of approximately 4400 to 5200 feet and slopes of 4 to 8 percent. Single-leaf pinyon pine, Muller’s oak, and California juniper dominate the overstory, and blackbrush is the dominant low shrub. The Xeric soil moisture regime and deep soils interspersed with bedrock outcrops contribute to high shrub diversity for a blackbrush dominated site.
Single-leaf pinyon pine is a long-lived, slow-growing tree that is a widespread dominant of Mojave Desert woodlands on shallow soils on soils with xeric moisture regimes, and where precipitation is 8 to 18 inches (Zouhar 2001, Minnich 2007). Muller’s oak is a long-lived, deep-rooted evergreen shrub that occurs in areas of higher rainfall such as the western margins of Mojave and Colorado Desert mountains and into the eastern slopes of the Peninsular Ranges in the California Floristic Province (Baldwin et al. 2002). California juniper is a long-lived small tree or large shrub that is a co-dominant in arid woodlands and scrub throughout California, achieving maximum dominance where mean annual precipitation is above 9.5 inches (Rhode 2002). Blackbrush is a dominant shrub in the mid-elevation zones of the Mojave and Great Basin Deserts. Extreme drought-tolerance coupled with extreme longevity allows blackbrush to achieve community dominance over shorter-lived species, provided there are long periods of time without disturbance (Pendleton and Meyer 2004). Thus, blackbrush communities are usually characterized by high blackbrush dominance, with a minor contribution of secondary shrubs (Brooks and Matchett 2003, Brooks et al. 2007).
The deep soils and additional runoff from bedrock outcrops on this site increase the availability of water, which increases the competitive ability of single-leaf pinyon pine, Muller’s oak and California juniper relative to blackbrush, and increase the suitability of this site to species like desert bitterbrush (Purshia glandulosa) and desert almond (Prunus fasciculata).
Disturbance Dynamics
Invasion by non-native species, drought and insect attack, and fire are the primary disturbances affecting this ecological site.
Non-native annual grasses (red brome [Bromus rubens] and cheatgrass [Bromus tectorum]) have become naturalized throughout the Mojave Desert over the past century (Rickard and Beatley 1965, D'Antonio and Vitousek 1992, Brooks 1999, Reid et al. 2006, Norton et al. 2007). The abundance and biomass of these grasses is highest on sandy soils with higher soil moisture availability, and where nitrogen deposition from air pollution from adjacent urban areas is high (Rao and Allen 2010, Rao et al. 2010). This ecological site receives relatively heavy nitrogen deposition due to its proximity to the greater Los Angeles area (Allen et al. 2009), and the deep sandy soils and relatively high precipitation regime of this site make it particularly susceptible to invasion by cheatgrass and red brome.
Invasion by non-native annual grasses has increased the flammability of Mojave Desert shrub communities by providing a continuous fine fuel layer between widely spaced shrubs (Brown and Minnich 1986, Brooks 1999, Brooks et al. 2004, Rao and Allen 2010, Rao et al. 2010). After fire, these communities appear to be more susceptible to invasion by exotic grasses, leading to a grass-fire cycle (D'Antonio and Vitousek 1992). The historic fire return interval in southern California pinyon-juniper woodlands is approximately 480 years, and in some stands there is no evidence of widespread fire (Minnich 2007, Romme et al. 2009). Similarly, fire in blackbrush shrublands was historically very rare (Brooks et al. 2007). Most desert community dominants are not adapted to fire, are slow-growing and long-lived, and require very long periods of time without disturbance to re-establish dominance. Thus an increase in fire frequency threatens to transition these communities to a permanently altered state.
Drought, and interactions with insect attack and disease are the most important drivers of community dynamics within pinyon-juniper woodlands (Minnich 2007, Romme et al. 2009). Pinyon-juniper woodlands are highly susceptible to drought, with widespread mortality and increased susceptibility to insect attack during drought (Shaw 2006, Minnich 2007, Romme et al. 2009). Trees in southern California close to urban centers may be especially at risk because increased nitrogen deposition from air pollution further increases susceptibility of pinyon pine to pathogens (Jones et al. 2004, Allen et al. 2010). Pinyon-juniper woodlands in the southwest experienced unprecedented drought and insect-induced mortality in the early 2000’s, with higher mortality in lower elevation stands, and a significant increase in standing dead pinyon pines (Shaw 2006). Cone-bearing trees (> 35 years of age) are more likely to die during drought, so older stands may be more severely drought-affected (Romme et al. 2009). This ecological site occupies relatively low elevations and receives relatively heavy nitrogen deposition due to its proximity to the greater Los Angeles area (Allen et al. 2009); thus, this ecological site may be especially vulnerable to the effects of drought.
State and transition model
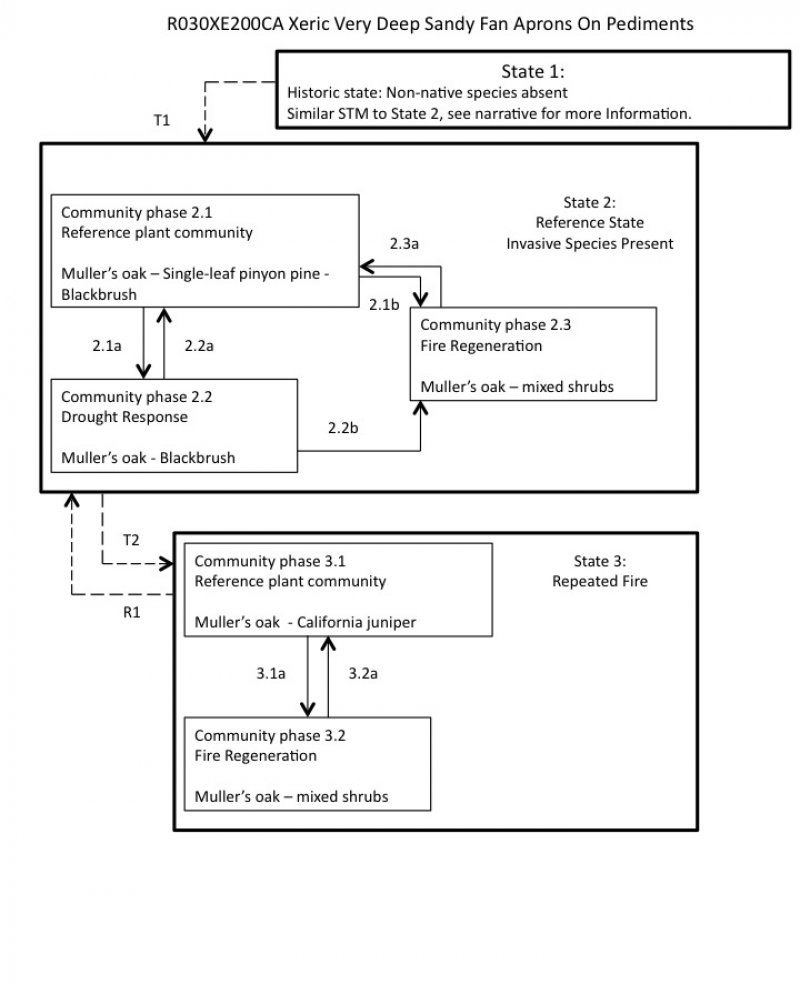
Figure 4. R030XE200CA
More interactive model formats are also available.
View Interactive Models
More interactive model formats are also available.
View Interactive Models
Click on state and transition labels to scroll to the respective text
Ecosystem states
State 2 submodel, plant communities
State 3 submodel, plant communities
State 1
Historic State
State 1 represents the historic range of variability for this ecological site. This state no longer exists due to the ubiquitous naturalization of non-native species in the Mojave Desert. Periodic drought and very rare fire were the natural disturbances influencing this ecological site. Data for this State does not exist, but it would have been similar to State 2, except with only native species present. See State 2 narrative for more detailed information.
State 2
Reference State
State 2 represents the current range of variability for this site. Non-native annuals, including red brome and cheatgrass are naturalized in this plant community. Their abundance varies with precipitation, but they are at least sparsely present (as current year’s growth or present in the soil seedbank).
Community 2.1
Reference plant community

Figure 5. Community Phase 2.1
This community has developed over time in the absence of fire.Single-leaf pinyon pine, Muller’s oak, and California juniper dominate the overstory, and blackbrush is the dominant low shrub. Desert bitterbrush (Purshia glandulosa), which is associated with the xeric precipitation regime and deep, coarse soils (Zlatnik 1999), is an important secondary shrub. Desert almond (Prunus fasciculata), Mojave yucca (Yucca schidigera), Joshua tree (Yucca brevifolia), and Nevada jointfir (Ephedra nevadensis) are also important secondary species. Native perennial bunchgrasses are sparsely present, and include desert needlegrass (Achnatherum speciosum), Indian ricegrass (Achnatherum hymenoides), and Sandberg bluegrass (Poa secunda). Native winter annual forbs may be abundant, and common species include bristly fiddleneck (Amsinckia tessellata), distant phacelia (Phacelia distans), and Great Basin langloisia (Langloisia setosissima). Non-native annual grasses may also be abundant, and include cheatgrass and red brome.
Figure 6. Annual production by plant type (representative values) or group (midpoint values)
Table 5. Annual production by plant type
| Plant type | Low (kg/hectare) |
Representative value (kg/hectare) |
High (kg/hectare) |
|---|---|---|---|
| Grass/Grasslike | – | 157 | 213 |
| Shrub/Vine | 101 | 157 | 213 |
| Tree | 90 | 135 | 179 |
| Forb | 1 | 25 | 34 |
| Total | 192 | 474 | 639 |
Community 2.2
Drought response
This community phase develops after prolonged or severe drought, and is characterized by mortality of single-leaf pinyon pine and overall declines in cover, with mortality also occurring in perennial grasses and short-lived shrubs. Pinyon pine mortality is more severe in older trees, and the prevalence of large, standing dead trees increases. Mortality may occur as a direct impact from water stress, or indirectly, from attack by Pinyon Ips beetle (Ips confuses) of drought-stressed trees. Muller’s oak has extremely high drought-tolerance, and since it is shade-intolerant, it may increase in response to the mortality of pine. California juniper is extremely drought-tolerant (Willson et al. 2008), with low rates of mortality during drought relative to pinyon pine (Breshears et al. 2005, Shaw 2006, Breshears et al. 2008, Floyd et al. 2009, Allen et al. 2010). Mortality rates of juniper species in response to drought in the early 2000’s range from 2 to 26% (Breshears et al. 2005, Mueller et al. 2005, Floyd et al. 2009). Blackbrush has very low mortality in response to drought, but may exhibit branch-pruning (Webb et al. 2003). Pinyon pine is slow-growing, recruitment episodic, and seedling establishment is most successful under cover of nurse plants or rocks (Chambers et al. 1999, Pearson and Theimer 2004). Further, pinyon is dependent on seed-caching rodents for dispersal to safe sites (Chambers et al. 1999), and rodent populations also decline during drought (Beatley 1969). With reduced seed production due to mortality, fewer dispersers, fewer safe sites for establishment and slow growth rates of the individuals that do establish, the effects of drought-induced pine mortality are long-lasting. Dead standing trees persist, and their replacement will take decades.
Community 2.3
Fire regeneration community

Figure 7. Community Phase 2.3
This community phase is characterized by the loss of pinyon pine, California juniper and blackbrush, with dominance by Muller’s oak. Pinyon pine, California juniper and blackbrush are generally killed by even moderate fire intensity (Romme et al. 2009). Muller’s oak is a vigorous resprouter, and will begin recovery in the first year after fire. Desert bitterbrush, desert almond, and Mojave yucca are also vigorous resprouters, and increase in importance in the fire regeneration community. The perennial grasses found in this site resprout and regenerate from seed within the first year after fire, thus increasing in response to fire (Sawyer et al. 2009). Threadleaf snakeweed (Gutierrezia microcephala) can rapidly recolonize from seed. Native and non-native annuals are abundant in the post-fire community, given adequate precipitation. As tall shrub cover increases, shade-dependent seedlings of California juniper and Pinyon pine begin to establish. Tree establishment may also occur in the shelter of boulders and rock outcrops (Pearson and Theimer 2004). California juniper re-establishes faster than pinyon pine (Tausch and West 1988). After fifty years with no disturbance, pine cover will begin to dominate, shading out shorter-lived shrubs, and mature woodlands re-establish at 100 to 150 years post-fire (Wangler and Minnich 1996). With favorable climatic conditions and nearby seed sources available in unburned pockets, blackbrush re-colonizes continuous soil patches. After long periods of time (> 100 years) with no disturbance, blackbrush gradually replaces shorter-lived species and regains dominance (Vasek 1983, Abella 2009, Vamstad 2009). This community is an at-risk phase. High biomass of non-native annual grasses after fire increases the susceptibility of this site to repeat burning. With frequent burning the fire return interval is too short to allow re-establishment of blackbrush, pinyon pine, and California juniper, and the site will transition to an altered state. The data presented for this community phase represent an example of vegetation recovery at four years after the West Complex Fire in Joshua Tree National Park. Trajectories of vegetation recovery in blackbrush shrub communities are highly divergent, and this divergence may persist for decades (Vamstad 2009, Engel and Abella 2011); thus this data is intended to be viewed as one potential trajectory.
Figure 8. Annual production by plant type (representative values) or group (midpoint values)
Table 6. Annual production by plant type
| Plant type | Low (kg/hectare) |
Representative value (kg/hectare) |
High (kg/hectare) |
|---|---|---|---|
| Grass/Grasslike | 3 | 157 | 213 |
| Forb | 4 | 84 | 118 |
| Shrub/Vine | 67 | 84 | 101 |
| Tree | – | – | 22 |
| Total | 74 | 325 | 454 |
Pathway 2.1a
Community 2.1 to 2.2
This pathway occurs with severe or prolonged drought.
Pathway 2.2a
Community 2.2 to 2.1
This pathway occurs with time, and a return to average or above average climatic conditions.
Pathway 2.3a
Community 2.3 to 2.1
This pathway occurs with time without fire.
State 3
Repeated fire
This state develops with a frequent fire return interval in the reference state. This state has been significantly altered from the natural range of variability found in States 1 and 2. Pinyon pine, California juniper, and blackbrush are largely lost, and Muller’s oak, non-native annual grasses, native sub-shrubs, perennial grasses, and short-lived shrubs dominate the community.
Community 3.1
Reference plant community
This reference plant community for State 3 is dominated by Muller’s oak, with desert bitterbrush, desert almond, short-lived shrubs, perennial grasses and native and non-native annual forbs and grasses.
Community 3.2
Fire regeneration community
The fire regeneration community is dominated by non-native annual species including cheatgrass, and red brome, and native forbs, including bristly fiddleneck, chia and pincushion flower (many other native forbs could also be present). Desert needlegrass, Indian ricegrass and Sandberg bluegrass may be abundant. There is a sparse cover of resprouting shrubs including Muller’s oak, desert bitterbrush and desert almond. This community is at high-risk of repeat burning due to high fine fuel cover. This community is also susceptible to wind and water erosion, due to the loss of stabilizing shrub cover (Bull 1997).
Pathway 3.1a
Community 3.1 to 3.2
This pathway occurs with moderate to severe fire.
Pathway 3.2a
Community 3.2 to 3.1
This pathway occurs with time without fire.
Transition 1
State 1 to 2
This transition occurred with the naturalization of non-native species in this ecological site. Non-native species were introduced with settlement of the Mojave Desert region in the 1860s.
Transition 3
State 2 to 3
This transition occurs if the fire return interval is less than 75 years.
Restoration pathway 1
State 3 to 2
Burned, blackbrush lost - State 3 This state is characterized by the loss of blackbrush from the plant community due to severe or recurrent fire. Data does not exist for this state, and community phase composition is based on literature review. The dynamics for this state are assumed to be similar to that described in State 2, except that blackbrush is no longer part of the plant community. Blackbrush lost Reference plant community - Community Phase 3.1 The reference plant community for this state is dominated by burrobush. Mojave yucca, Nevada ephedra, and eastern Mojave buckwheat are important secondary species. Other shrub species, subshrubs and annuals are likely to remain as described in Community phase 2.1. Community Phase Pathway 3.1a This pathway occurs in response to prolonged or severe drought. Community Phase Pathway 3.1b This pathway occurs in response to moderate or severe fire. Restoration Pathway - 1 Restoration of communities severely altered by repeat fire at the landscape scale is difficult. Methods may include aerial seeding of early native colonizers such as desert globemallow, desert needlegrass, Indian ricegrass, and threadleaf snakeweed. Increased native cover may help to reduce non-native plant invasion, helps to stabilize soils, provides a source of food and cover for wildlife, and provides microsites that facilitate California juniper, singleleaf pinyon pine, and blackbrush establishment. However, the amount of seed required for success is often prohibitive. Large-scale planting of both early colonizers and community dominants tends to be more successful in terms of plant survival, especially if outplants receive supplemental watering during the first two years. Blackbrush is difficult to cultivate for outplanting due to susceptibility to fungal pathogens in greenhouse environment. Pre-emergent herbicides (Plateau) have been used in the year immediately post-fire to attempt to inhibit or reduce brome invasion. How successful this is on a landscape scale, and the non-target effects have not yet been determined.
Additional community tables
Table 7. Community 2.1 plant community composition
| Group | Common name | Symbol | Scientific name | Annual production (kg/hectare) | Foliar cover (%) | |
|---|---|---|---|---|---|---|
|
Tree
|
||||||
| 1 | Trees | 90–179 | ||||
| California juniper | JUCA7 | Juniperus californica | 56–135 | 3–8 | ||
| singleleaf pinyon | PIMO | Pinus monophylla | 22–45 | 2–4 | ||
| Joshua tree | YUBR | Yucca brevifolia | 1–22 | 0–2 | ||
|
Shrub/Vine
|
||||||
| 2 | Shrubs | 101–213 | ||||
| Muller oak | QUCO7 | Quercus cornelius-mulleri | 56–112 | 2–5 | ||
| blackbrush | CORA | Coleogyne ramosissima | 29–58 | 5–20 | ||
| desert bitterbrush | PUGL2 | Purshia glandulosa | 19–38 | 0–3 | ||
| Nevada jointfir | EPNE | Ephedra nevadensis | 0–12 | 0–4 | ||
| desert almond | PRFA | Prunus fasciculata | 0–11 | 0–4 | ||
| Mojave yucca | YUSC2 | Yucca schidigera | 0–11 | 0–4 | ||
| Mexican bladdersage | SAME | Salazaria mexicana | 0–1 | 0–1 | ||
| Wiggins' cholla | CYEC3 | Cylindropuntia echinocarpa | 0–1 | 0–1 | ||
| kingcup cactus | ECTR | Echinocereus triglochidiatus | 0–1 | 0–1 | ||
| Eastern Mojave buckwheat | ERFA2 | Eriogonum fasciculatum | 0–1 | 0–1 | ||
| threadleaf snakeweed | GUMI | Gutierrezia microcephala | 0–1 | 0–1 | ||
| beavertail pricklypear | OPBA2 | Opuntia basilaris | 0–1 | 0–1 | ||
|
Forb
|
||||||
| 3 | Native forbs | 1–34 | ||||
| Great Basin langloisia | LASE3 | Langloisia setosissima | 0–34 | 0–2 | ||
| bristly fiddleneck | AMTE3 | Amsinckia tessellata | 0–11 | 0–1 | ||
| distant phacelia | PHDI | Phacelia distans | 0–11 | 0–1 | ||
| Parry's wirelettuce | STPA3 | Stephanomeria parryi | 0–6 | 0–1 | ||
| chia | SACO6 | Salvia columbariae | 0–1 | 0–1 | ||
| desert globemallow | SPAM2 | Sphaeralcea ambigua | 0–1 | 0–1 | ||
| woolly easterbonnets | ANWA | Antheropeas wallacei | 0–1 | 0–1 | ||
| bluedicks | DICA14 | Dichelostemma capitatum | 0–1 | 0–1 | ||
| buckwheat | ERIOG | Eriogonum | 0–1 | 0–1 | ||
| whitedaisy tidytips | LAGL5 | Layia glandulosa | 0–1 | 0–1 | ||
| golden linanthus | LEAUA3 | Leptosiphon aureus ssp. aureus | 0–1 | 0–1 | ||
| Mojave desertparsley | LOMO | Lomatium mohavense | 0–1 | 0–1 | ||
| miniature lupine | LUBI | Lupinus bicolor | 0–1 | 0–1 | ||
| blazingstar | MENTZ | Mentzelia | 0–1 | 0–1 | ||
| desert wishbone-bush | MILA6 | Mirabilis laevis | 0–1 | 0–1 | ||
| 4 | Non-native annual forbs | 0–1 | ||||
| redstem stork's bill | ERCI6 | Erodium cicutarium | 0–1 | 0–1 | ||
|
Grass/Grasslike
|
||||||
| 5 | Perennial grasses | 0–17 | ||||
| desert needlegrass | ACSP12 | Achnatherum speciosum | 0–11 | 0–1 | ||
| Sandberg bluegrass | POSE | Poa secunda | 0–1 | 0–1 | ||
| Indian ricegrass | ACHY | Achnatherum hymenoides | 0–1 | 0–1 | ||
| 6 | Non-native annual grasses | 0–213 | ||||
| compact brome | BRMA3 | Bromus madritensis | 0–157 | 0–5 | ||
| cheatgrass | BRTE | Bromus tectorum | 0–146 | 0–5 | ||
Table 8. Community 2.3 plant community composition
| Group | Common name | Symbol | Scientific name | Annual production (kg/hectare) | Foliar cover (%) | |
|---|---|---|---|---|---|---|
|
Tree
|
||||||
| 1 | Trees | 0–22 | ||||
| Joshua tree | YUBR | Yucca brevifolia | 0–22 | 0–2 | ||
| California juniper | JUCA7 | Juniperus californica | 0–6 | 0–1 | ||
|
Shrub/Vine
|
||||||
| 2 | Native shrubs | 67–101 | ||||
| Muller oak | QUCO7 | Quercus cornelius-mulleri | 0–84 | 0–3 | ||
| desert almond | PRFA | Prunus fasciculata | 0–28 | 0–7 | ||
| desert bitterbrush | PUGL2 | Purshia glandulosa | 0–26 | 0–2 | ||
| Nevada jointfir | EPNE | Ephedra nevadensis | 0–13 | 0–2 | ||
| Mojave yucca | YUSC2 | Yucca schidigera | 0–11 | 0–4 | ||
| threadleaf snakeweed | GUMI | Gutierrezia microcephala | 0–6 | 0–1 | ||
| blackbrush | CORA | Coleogyne ramosissima | 0–2 | 0–1 | ||
|
Grass/Grasslike
|
||||||
| 3 | Perennial Grasses | 0–4 | ||||
| desert needlegrass | ACSP12 | Achnatherum speciosum | 0–3 | 0–1 | ||
| Indian ricegrass | ACHY | Achnatherum hymenoides | 0–1 | 0–1 | ||
| 6 | Non-native annual grasses | 0–157 | ||||
| cheatgrass | BRTE | Bromus tectorum | 0–146 | 0–20 | ||
| red brome | BRRU2 | Bromus rubens | 0–11 | 0–1 | ||
|
Forb
|
||||||
| 4 | Native forbs | 4–118 | ||||
| smooth desertdandelion | MAGL3 | Malacothrix glabrata | 0–56 | 0–2 | ||
| Great Basin langloisia | LASE3 | Langloisia setosissima | 0–34 | 0–2 | ||
| small wirelettuce | STEX | Stephanomeria exigua | 0–22 | 0–3 | ||
| bristly fiddleneck | AMTE3 | Amsinckia tessellata | 0–11 | 0–1 | ||
| desert globemallow | SPAM2 | Sphaeralcea ambigua | 0–6 | 0–1 | ||
| Parry's wirelettuce | STPA3 | Stephanomeria parryi | 0–6 | 0–1 | ||
Interpretations
Animal community
This ecological site provides important habitat for birds, reptiles and mammals due to its structural diversity and pinyon woodlands.
The following reptiles and mammals are likely to be found within this ecological site (based on habitat preferences).
REPTILES
Lizards:
Desert banded Gecko (Coleonyx variegatus variegatus)
Northern desert iguana (Dipsosaurus dorsalis dorsalis)
Mojave collared lizard (Crotaphytus bicinctores)
Western chuckwalla (Sauromalus aster obesus)
Yellow-backed spiny lizard (Sceloporus magister uniformus)
Great Basin fence lizard (Sceloporus biseriatus longipes)
Western brush lizard (Urosaurus graciosus graciosus)
Desert side-blotched lizard (Uta stansburiana stejnegeri)
Great Basin Whiptail (Aspidoscelis tigris tigris)
Snakes:
Southwestern blind snake (Leptotyphlops humilis humilis)
Desert rosy boa (Lichanura trivirgata gracia)
Mojave glossy snake (Arizona occidentalis candida) California kingsnake (Lampropeltis getula californae)
Red coachwhip (Masticophis flagellum piceus)
Desert night snake (Hypsiglena torquata deserticola)
California kingsnake (Lampropeltis getula californae)
California striped racer (Masticophis lateralis lateralis)
Western leaf-nosed snake (Phyllorynchus decurtatus perkinsi)
Great Basin gopher snake (Pituophis catenifer deserticola)
Western long-nosed snake (Rhinocheilus lecontei lecontei)
Smith’s black-headed snake (Tantilla hobartsmithi)
California lyre snake (Trimorphodon biscutatus vandenburghi)
Southwestern speckled rattlesnake (Crotalus mitchelli Pyrrhus)
Red diamond rattlesnake (Crotalus ruber ruber)
Southern pacific rattlesnake (Crotalus helleri)
MAMMALS
Western spotted skunk (Spilogale gracilis gracilis)
Long-tailed weasel (Mustela latirosta)
California desert bat (Myotis californicus stephensi)
Western pipistrelle (Pipistrellus hesperus hesperus)
Desert big brown bat (Eptesicus fuscus pallidus)
Desert long-legged bat (Myotis volans interior)
Northern fringed bat (Myotis thysanodes thysanodes)
Spotted bat (Euderma maculatum)
Western mastiff bat (Eumops perotis)
Hoary bat (Lasiurus cinereus cinereus)
Pallid bat (Antrozous pallidus minor)
Desert coyote (Canis macrotis arsipus)
Common gray fox (Urocyon cinereoargenteus scottii)
California mountain lion (Felis concolor californica)
Desert bobcat (Lynx rufus baileyi)
California ringtail (Bassariscus astutus ocatvus)
Southern mule deer (Odoceileus hemionus fuliginatus)
Desert bighorn sheep (Ovis canadensis nelson)
Southern Desert cottontail (Sylvilagus audobonii arizonae)
Dusky chipmonk (Tamias obscurus davisi)
Whitetail antelope squirrel (Ammospermphilus leucurus leucurus)
Western Mojave ground squirrel (Spermophilus beecheyi parvulus)
Long-tailed pocket mouse (Chaetodipus mojavensis)
Merriam's kangaroo rat (Dipodomys deserti)
Desert wood rat (Neotoma fuscipes simplex)
Eastern dusky-footed wood rat (Neotoma fuscipes simplex)
White-throated wood rat (Neotoma albigula venusta)
Desert canyon mouse (Peromyscus crinitus stephensi)
Southern brush mouse (Peromyscus boylii rowleyi)
Sonoran deer mouse (Peromyscus maniculatus sonoriensis)
Southern California pinyon mouse (Peromyscus truei chlorus)
Desert grasshopper mouse (Onychomys torridus pulcher)
Desert shrew
Recreational uses
This ecological site occurs in remote, steep terrain with limited access. As such, it has important wilderness values. It may be used for cross-country hiking, solitude, botanizing, and aesthetic enjoyment.
Wood products
Wood of single-leaf pinyon may be used for fuel wood and fence posts, particle and cement board (Zouhar 2001). It is not suitable for lumber because of its small size and irregular growth pattern (Zouhar 2001).
California juniper is a poor source of lumber because of low volume and multi-stemmed growth form. However, early ranchers used juniper for fenceposts, and it is used for fuel and as Christmas trees (Cope 1992).
Other products
Single-leaf pinyon pine is very important for many Native American tribes. The melted gum is used to bind and heal cuts, prevent sunburn, to stop menstruation, for muscle soreness, diarhea, rheumatism, colds, and nausea, among others. Pinyon nuts are an important food source, and were the staple food source in the past for many tribes. The pinyon seeds were one of the few foods given to babies as an alternative food source by the Cahuilla. Needles are used to make baskets, and were used as a spice to flavor meats. Wood and bark were used in house construction, and the pitch was used for water-proofing. http://herb.umd.umich.edu/herb/search.pl?searchstring=Pinus%20monophylla).
Supporting information
Inventory data references
The following NRCS plots were used to describe this ecological site:
Community Phase 2.1:
1249800639 (Type location)
1251503837
1249806A
Community Phase 2.3:
1249806705
1249806932
Type locality
| Location 1: San Bernardino County, CA | |
|---|---|
| UTM zone | N |
| UTM northing | 563348 |
| UTM easting | 3765774 |
| General legal description | The type location is approximately 0.2 miles east of Covington Flats Crossover Road, and 1.3 miles southwest of the intersection of Covington Flats Crossover Road and Lower Covington Flat Road in Joshua Tree National Park. |
Other references
Allen, E. B., L. E. Rao, R. J. Steers, A. Bytnerowicz, and M. E. Fenn. 2009. Impacts of atmospheric nitrogen deposition on vegetation and soils at Joshua Tree National Park. Pages 78-100 in R. H. Webb, L. F. Fenstermaker, J. S. Heaton, D. L. Hughson, E. V. McDonald, and D. M. Miller, editors. The Mojave Desert. University of Nevada Press, Reno, Nevada.
Allen, M. F., E. B. Allen, J. L. Lansing, K. S. Pregitzer, R. L. Hendrick, R. W. Ruess, and S. L. Collins. 2010. Responses to chronic N fertilization of ectomycorrhizal pinon but not arbuscular mycorrhizal juniper in a pinon-juniper woodland. Journal of Arid Environments.
Baldwin, B. G., S. Boyd, B. J. Ertter, R. W. Patterson, T. J. Rosatti, and D. H. Wilken. 2002. The Jepson Desert Manual. University of California Press, Berkeley and Los Angeles, California.
Beatley, J. C. 1969. Dependence of desert rodents on winter annuals and precipitation. Ecology 50:721-724.
Breshears, D. D., N. S. Cobb, P. M. Rich, K. P. Price, C. D. Allen, R. G. Balice, W. H. Romme, J. H. Kastens, M. L. Floyd, J. Belnap, J. J. Anderson, O. B. Myers, and C. W. Meyer. 2005. Regional vegetation die-off in response to global-change-type drought. Proceedings of the National Academy of Sciences of the United States of America 102:15144-15148.
Breshears, D. D., O. B. Myers, C. W. Meyer, F. J. Barnes, C. B. Zou, C. D. Allen, N. G. McDowell, and W. T. Pockman. 2008. Tree die-off in response to global change-type drought: mortality insights from a decade of plant-water potential measurements. Frontiers in Ecology.
Brooks, M. L. 1999. Habitat invasibility and dominance by alien annual plants in the western Mojave Desert. Biological Invasions 1:325-337.
Brooks, M. L. and J. R. Matchett. 2003. Plant community patterns in unburned and burned blackbrush (Coleogyne ramosissima Torr.) shrublands in the Mojave Desert. Western North American Naturalist 63:283-298.
Brooks, M. L., C. M. D'Antonio, D. M. Richardson, J. B. Grace, J. E. Keeley, J. M. DiTomaso, R. J. Hobbs, M. Pellant, and D. Pyke. 2004. Effects of invasive alien plants on fire regimes. Bioscience 54:677-689.
Brooks, M. L., T. C. Esque, and T. Duck. 2007. Creosotebush, blackbrush, and interior chaparral shrublands. RMRS-GTR-202.
Brown, D. E. and R. A. Minnich. 1986. Fire and Changes in Creosote Bush Scrub of the Western Sonoran Desert, California. American Midland Naturalist 116:411-422.
Chambers, J. C., S. B. V. Wall, and E. W. Schupp. 1999. Seed and seedling ecology of piñon and juniper seeds in the pygmy woodlands of western North America. The Botanical Review 65:2-36.
Cope, Amy B. 1992. Juniperus californica. In: Fire Effects Information System, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer).Available: http://www.fs.fed.us/database/feis/ [2012, April 2].
D'Antonio, C. M. and P. M. Vitousek. 1992. Biological invasions by exotic grasses, the grass/fire cycle, and global change. Annual Review of Ecology and Systematics 23:63-87.
Engel, E. C. and S. R. Abella. 2011. Vegetation recovery in a desert landscape after wildfires: influences of community type, time since fire and contingency effects. Journal of Applied Ecology 48:1401-1410.
Floyd, M. L., M. Clifford, N. S. Cobb, D. Hanna, R. Delph, P. Ford, and D. Turner. 2009. Relationship of stand characteristics to drought-induced mortality in three southwestern pinon-juniper woodlands. Ecological Applications 19:1223-1230.
Jones, M. E., T. D. Paine, M. E. Fenn, and M. A. Poth. 2004. Influence of ozone and nitrogen deposition on bark beetle activity under drought conditions. Forest Ecology and Management 200:67-76.
Minnich, R. A. 2003. Fire and dynamics of temperature desert woodlands in Joshua Tree National Park. Contract, Joshua Tree National Park.
Minnich, R. A. 2007. Southern California conifer forests. Pages 502-539.
Mueller, R. C., C. M. Scudder, M. E. Porter, R. T. T. III, C. A. Gehring, and T. G. Whitham. 2005. Differential tree mortality in response to severe drought: evidence for long-term vegetation shifts. Journal of Ecology.
Norton, J. B., T. A. Monaco, and U. Norton. 2007. Mediterranean annual grasses in western North America: kids in a candy store. Plant Soil 298:1-5.
Pearson, K. M. and T. C. Theimer. 2004. Seed-caching responses to substrate and rock cover by two Peromyscus species: implications for pinyon pine establishment. Oecologia 141:76-83.
Rao, L. E. and E. B. Allen. 2010. Combined effects of precipitation and nitrogen deposition on native and invasive winter annual production in California deserts. Oecologia 162:1035-1046.
Rao, L. E., E. B. Allen, and T. M. Meixner. 2010. Risk-based determination of critical nitrogen deposition loads for fire spread in southern California deserts. Ecological Applications 20:1320-1335.
Reid, C. R., S. Goodrich, and J. E. Bowns. 2006. Cheatgrass and red brome: history and biology of two invaders. Pages 27-32 in Shrublands under fire: disturbance and recovery in a changing world. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Cedar City, Utah.
Rickard, W. H. and J. C. Beatley. 1965. Canopy-coverage of the desert shrub vegetation mosaic of the Nevada test site. Ecology 46:524-529.
Rhode, D. 2002. Early Holocene juniper woodland and chaparral taxa in the central Baja California Peninsula, Mexico. Quaternary Research 57:102-108.
Romme, W. H., C. D. Allen, J. D. Bailey, W. L. Baker, B. T. Bestelmeyer, P. M. Brown, K. S. Eisenhart, M. L. Floyd, D. W. Huffman, B. F. Jacobs, R. F. Miller, E. H. Muldavin, T. W. Swetnam, R. J. Tausch, and P. J. Weisberg. 2009. Historical and Modern Disturbance Regimes, Stand Structures, and Landscape Dynamics in Pinon–Juniper Vegetation of the Western United States. Rangeland Ecological Management 62:203-222.
Sawyer, J. O., T. Keeler-Woolf, and J. M. Evans. 2009. A manual of California vegetation. 2nd edition. California Native Plant Society, Sacramento, California.
Shaw, J. D. 2006. Population-wide changes in Pinyon-Juniper woodlands caused by drought in the American Southwest: effects on structure, composition, and distribution. Page 8 in IUFRO Landscape Ecology Conference, Locorontondo, Bari (Italy).
Tausch, R. J. and N. E. West. 1988. Differential establishment of pinyon and juniper following fire. The American Midland Naturalist 119:174-184.
Vamstad, M. S. 2009. Effects of fire on vegetation and small mammal communities in a Mojave Desert Joshua tree woodland. M.S. Unversity of California, Riverside, Riverside, Ca.
Vasek, F. C. 1983. Plant succession in the Mojave Desert. Crossosoma 9:1-23.
Wangler, M. J. and R. A. Minnich. 1996. Fire and succession in pinyon-junipe woodlands of the San Bernardino Mountains, California. Madroño 43:493-514.
Webb, R. H., M. B. Muroy, T. C. Esque, D. E. Boyer, L. A. DeFalco, D. F. Haines, D. Oldershaw, S. J. Scoles, K. A. Thomas, J. B. Blainey, and P. A. Medica. 2003. Perennial vegetation data from permanent plots on the Nevada Test Site, Nye County, Nevada. U.S. Geological Society, Tucson, AZ.
Willson, C. J., P. S. Manos, and R. B. Jackson. 2008. Hydraulic traits are influenced by phylogenetic history in the drought-resistant, invasive genus Juniperus (Cupressaceae). American Journal of Botany 95:299-314.
Zlatnik, Elena. 1999. Purshia glandulosa. In: Fire Effects Information System, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer). Available: http://www.fs.fed.us/database/feis/ [2012, February 6].
Zouhar, Kristin L. 2001. Pinus monophylla. In: Fire Effects Information System, [Online]. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer). Available: http://www.fs.fed.us/database/feis/ [2012, January 16].
Contributors
Allison Tokunaga
Alice Lee Miller
Approval
Kendra Moseley, 10/21/2024
Rangeland health reference sheet
Interpreting Indicators of Rangeland Health is a qualitative assessment protocol used to determine ecosystem condition based on benchmark characteristics described in the Reference Sheet. A suite of 17 (or more) indicators are typically considered in an assessment. The ecological site(s) representative of an assessment location must be known prior to applying the protocol and must be verified based on soils and climate. Current plant community cannot be used to identify the ecological site.
| Author(s)/participant(s) | |
|---|---|
| Contact for lead author | |
| Date | 12/22/2024 |
| Approved by | Kendra Moseley |
| Approval date | |
| Composition (Indicators 10 and 12) based on | Annual Production |
Indicators
-
Number and extent of rills:
-
Presence of water flow patterns:
-
Number and height of erosional pedestals or terracettes:
-
Bare ground from Ecological Site Description or other studies (rock, litter, lichen, moss, plant canopy are not bare ground):
-
Number of gullies and erosion associated with gullies:
-
Extent of wind scoured, blowouts and/or depositional areas:
-
Amount of litter movement (describe size and distance expected to travel):
-
Soil surface (top few mm) resistance to erosion (stability values are averages - most sites will show a range of values):
-
Soil surface structure and SOM content (include type of structure and A-horizon color and thickness):
-
Effect of community phase composition (relative proportion of different functional groups) and spatial distribution on infiltration and runoff:
-
Presence and thickness of compaction layer (usually none; describe soil profile features which may be mistaken for compaction on this site):
-
Functional/Structural Groups (list in order of descending dominance by above-ground annual-production or live foliar cover using symbols: >>, >, = to indicate much greater than, greater than, and equal to):
Dominant:
Sub-dominant:
Other:
Additional:
-
Amount of plant mortality and decadence (include which functional groups are expected to show mortality or decadence):
-
Average percent litter cover (%) and depth ( in):
-
Expected annual annual-production (this is TOTAL above-ground annual-production, not just forage annual-production):
-
Potential invasive (including noxious) species (native and non-native). List species which BOTH characterize degraded states and have the potential to become a dominant or co-dominant species on the ecological site if their future establishment and growth is not actively controlled by management interventions. Species that become dominant for only one to several years (e.g., short-term response to drought or wildfire) are not invasive plants. Note that unlike other indicators, we are describing what is NOT expected in the reference state for the ecological site:
-
Perennial plant reproductive capability:
Print Options
Sections
Font
Other
The Ecosystem Dynamics Interpretive Tool is an information system framework developed by the USDA-ARS Jornada Experimental Range, USDA Natural Resources Conservation Service, and New Mexico State University.
Click on box and path labels to scroll to the respective text.