Ecological site group R023XY905NV
Loamy 10-20 PZ Mountain Big Sagebrush and Bluebunch Wheatgrass
Last updated: 06/03/2024
Accessed: 04/02/2026
Ecological site group description
Key Characteristics
- Site does not pond or flood
- Landform other than dunes
- Surface soils are not clayey
- Sites are shrub or grass dominated
- [Criteria]MAP >10"
- Soil is moderately deep or deeper
- Site is on south aspect of plateau landscapes
Provisional. A provisional ecological site description has undergone quality control and quality assurance review. It contains a working state and transition model and enough information to identify the ecological site.
Physiography
This group is on plateaus, fan piedmonts, and mountains throughout MLRA 23. The elevation is 4,500 to 7,500 feet.
Climate
The climate is classified as Cold Semi-Arid in the Koppen Classification System.
The area receives 10 to 20 inches of annual precipitation as snow in the winter and rain in spring and fall. Summers are generally dry.
The frost-free period is 40 to 110 days. The mean annual air temperature is 24 to 52 °F.
Soil features
The soils in this group are variable, but most are Mollisols at least 12 inches deep. They may be either mesic or frigid. The central concept is a loamy, moderately deep or deeper soil.
Some common soil series in this group include Hart Camp, Softscrabble, Sumine, Dosie, and Ashtre.
Vegetation dynamics
Ecological Dynamics and Disturbance Response:
An ecological site is the product of all the environmental factors responsible for its development. Each site has a set of key characteristics that influence its resilience to disturbance and resistance to invasives. According to Caudle et al. (2013), key characteristics include:
1. Climate factors such as precipitation and temperature.
2. Topographic characteristics such as aspect, slope, elevation, and landform.
3. Hydrologic processes such as infiltration and runoff.
4. Soil characteristics such as depth, texture, structure, and organic matter.
5. Plant communities and their functional groups and productivity.
6. Natural disturbance (fire, herbivory, etc.) regime.
Biotic factors that influence resilience include site productivity, species composition and structure, and population regulation and regeneration (Chambers et al., 2013).
The ecological sites in this group are dominated by deep-rooted, cool-season, perennial bunchgrasses and long-lived shrubs (at least 50 years old) with high root to shoot ratios. The dominant shrubs usually root to the full depth of the winter-spring soil moisture recharge, which ranges from 1.0 to over 3.0 meters (Dobrowolski et al.,1990). Root length of mature sagebrush plants was measured to a depth of 2 meters in alluvial soils in Utah (Richards & Caldwell, 1987). These shrubs have a flexible generalized root system with development of both deep taproots and laterals near the surface (Comstock & Ehleringer, 1992). Antelope bitterbrush (Purshia tridentata) is most common on soils with minimal restrictions that allow deep root penetration such as coarse-textured soil, or finer-textured soil with high stone content (Driscoll, 1964; Clements & Young, 2002).
The Great Basin sagebrush communities have high spatial and temporal variability in precipitation both among years and within growing seasons (MacMahon, 1980). Nutrient availability is typically low but increases with elevation and closely follows moisture availability. The invasibility of plant communities is often linked to resource availability. Disturbance changes resource uptake and increases nutrient availability, often to the benefit of non-native species; native species are often damaged and their ability to use resources is depressed for a time, but resource pools may increase from lack of use and/or the decomposition of dead plant material following disturbance (Whisenant, 1999; Miller et al., 2013). The invasion of sagebrush communities by cheatgrass (Bromus tectorum) has been linked to disturbances (fire, abusive grazing) that result in fluctuations in resources (Beckstead & Augspurger, 2004; Chambers et al., 2007; Johnson et al., 2011).
Periodic drought regularly influences sagebrush ecosystems, and drought duration and severity have increased throughout the 20th century in much of the Intermountain West. Major shifts away from historical precipitation patterns have the greatest potential to alter ecosystem function and productivity. Species composition and productivity can be altered by the timing of precipitation and water availability within the soil profile (Bates et al., 2006).
Native insect outbreaks are also important drivers of ecosystem dynamics in big sagebrush (Artemisia tridentata) communities. Climate influences the timing of insect outbreaks, especially outbreaks of a sagebrush defoliator called Aroga moth (Aroga websteri). Aroga moth infestations occurred in the Great Basin in the 1960s, the early 1970s, and have been ongoing in Nevada since 2004 (Bentz et al., 2008). Thousands of acres of big sagebrush have been impacted, with partial to complete die-off observed. The Aroga moth can partially or entirely kill individual plants or entire stands of big sagebrush (Furniss & Barr, 1975).
The dominant perennial bunchgrasses include bluebunch wheatgrass (Pseudoroegneria spicata), Thurber’s needlegrass (Achnatherum thurberianum), and basin wildrye (Leymus cinereus). These grass species generally have somewhat shallower root systems than the shrubs on these sites; root densities of these grasses are often as high as or higher than those of shrubs in the upper 0.5 meters of the soil profile. The general differences in root depth distributions between these grasses and shrubs result in resource partitioning in these shrub/grass systems.
Beardless wildrye (Leymus triticoides), also known as creeping wildrye, is a subdominant grass on this site. It is a cool-season, perennial, sod-forming grass that is strongly rhizomatous (Young-Mathews & Winslow, 2010). In a study of native grasses in California, beardless wildrye performed the best in terms of above-ground biomass and high resistance to invasion by non-native annuals (Lulow, 2006).
Where sites in this group are adjacent to juniper stands, there is potential for infilling by Utah juniper (Juniperus osteosperma) or western juniper (J. occidentalis). Without disturbance in these areas, juniper will eventually dominate the site and out-compete sagebrush for water and sunlight, causing a severe reduction of both the shrub and herbaceous understory (Miller & Tausch, 2001; Lett & Knapp, 2005). The potential for soil erosion increases as the woodland matures and the understory plant community cover declines (Pierson et al., 2010).
The ecological sites in this group have moderate to high resilience to disturbance and resistance to invasion. Resilience increases with elevation, northerly aspect, precipitation, and nutrient availability. Five possible states have been identified for the South Slope 12-16” ecological site. Differences in resilience to disturbance for the remaining ecological sites contained within this group are described at the end of this document.
Annual Invasive Grasses:
The species most likely to invade these sites are cheatgrass and medusahead (Taeniatherum). Both species are cool- season annual grasses that maintain an advantage over native plants in part because they are prolific seed producers, able to germinate in the autumn or spring, tolerant of grazing, and increase with frequent fire (Klemmedson & Smith, 1964; Miller et al., 1999). Medusahead and cheatgrass originated from Eurasia and both were first reported in North America in the late 1800s (Mack & Pyke, 1983; Furbush, 1953). Pellant and Hall (1994) found 3.3 million acres of public lands dominated by cheatgrass and suggested that another 76 million acres were susceptible to invasion by winter annuals including cheatgrass and medusahead. By 2003, medusahead occupied approximately 2.3 million acres in 17 western states (Rice, 2005). In the Intermountain West, the exponential increase in dominance by medusahead has largely been at the expense of cheatgrass (Harris, 1967; Hironaka, 1994). Medusahead matures 2 to 3 weeks later than cheatgrass (Harris, 1967). Recently, James et al. (2008) measured leaf biomass over the growing season and found that medusahead maintained vegetative growth later in the growing season than cheatgrass. Mangla et al. (2011) also found medusahead had a longer period of growth and more total biomass than cheatgrass and hypothesized this difference in relative growth rate may be due to the ability of medusahead to maintain water uptake as upper soils dry compared to co-occurring species, especially cheatgrass. Medusahead litter has a slow decomposition rate because of its high silica content, allowing it to accumulate over time and suppress competing vegetation (Bovey et al., 1961; Davies & Johnson, 2008). Harris (1967) reported medusahead roots have thicker cell walls compared to those of cheatgrass, allowing it to more effectively conduct water, even in very dry conditions.
Recent modeling and empirical work by Bradford and Lauenroth (2006) suggest that seasonal patterns of precipitation input and temperature are also key factors determining regional variation in the growth, seed production, and spread of invasive annual grasses. Collectively, the body of research suggests that the invasion and dominance of medusahead onto native grasslands and cheatgrass-infested grasslands will continue to increase in severity because conditions that favor native bunchgrasses or cheatgrass over medusahead are rare (Mangla et al. 2011). Medusahead replaces native vegetation and cheatgrass directly by competition and suppression. It replaces native vegetation indirectly by increasing fire frequency.
Methods to control medusahead and cheatgrass include herbicide, fire, grazing, and seeding of primarily non-native wheatgrasses. Mapping potential or current invasion vectors is a management method designed to increase the cost effectiveness of control methods. A study by Davies et al. (2013) found an increase in medusahead cover near roads. Cover was higher near animal trails than random transects, but the difference was less evident. This implies that vehicles and animals aid the spread of the weed; however, vehicles are the major vector of movement. Spraying with herbicide (Imazapic or Imazapic and glyphosate) and seeding with crested wheatgrass (Agropyron cristatum) and Sandberg bluegrass (Poa secunda) have been more successful at combating medusahead and cheatgrass than spraying alone (Sheley et al., 2012). Where native bunchgrasses are missing from the site, revegetation of medusahead- or cheatgrass-invaded rangelands has a higher likelihood of success when using introduced perennial bunchgrasses such as crested wheatgrass (Davies et al., 2015). Butler et al. (2011) tested four herbicides (Imazapic, Imazapic + glyphosate, rimsulfuron, and sulfometuron + Chlorsulfuron), using herbicide-only treatments for suppression of cheatgrass, medusahead, and ventenata (Ventenata dubia) within residual stands of native bunchgrass. Additionally, they tested the same four herbicides followed by seeding of six bunchgrasses (native and non-native) with varying success. Herbicide-only treatments appeared to remove competition for established bluebunch wheatgrass by providing 100 percent control of ventenata and medusahead and greater than 95 percent control of cheatgrass. However, caution in using these results is advised, as only one year of data was reported.
Prescribed fire has also been utilized in combination with the application of pre-emergent herbicide to control medusahead and cheatgrass (J. L. Vollmer & J. G. Vollmer, 2008). Mature medusahead or cheatgrass is very flammable and fire can be used to remove the thatch layer, consume standing vegetation, and even reduce seed levels. Furbush (1953) reported that timing a burn while the seeds were in the milk stage effectively reduced medusahead the following year. He further reported that adjacent unburned areas became a seed source for reinvasion the following year.
When considering the combination of pre-emergent herbicide and prescribed fire for invasive annual grass control, it is important to assess the tolerance of desirable brush species to the herbicide being applied. J. L. Vollmer and J. G. Vollmer (2008) tested the tolerance of mountain mahogany (Cercocarpus montanus), antelope bitterbrush, and multiple sagebrush species to three rates of Imazapic and the same rates with methylated seed oil as a surfactant. They found a cheatgrass control program in an antelope bitterbrush community should not exceed Imazapic at 8 ounces per acre with or without surfactant. Sagebrush, regardless of species or rate of application, was not affected. However, many environmental variables were not reported in this study and managers should install test plots before broad scale herbicide application is initiated.
Fire Ecology:
Pre-settlement fire return intervals in mountain big sagebrush (Artemisia tridentata ssp. vaseyana) communities varied from 15 to 25 years (Burkhardt & Tisdale, 1969; Houston, 1973; Miller et al., 2000). Mountain big sagebrush is killed by fire (Neuenschwander, 1980; Blaisdell et al., 1982), and does not resprout (Blaisdell, 1953). Post-fire regeneration originates from seed and will vary depending on site characteristics, seed sources, and fire characteristics. Mountain big sagebrush seedlings can grow rapidly and may reach reproductive maturity within 3 to 5 years (Bunting et al., 1987). Mountain big sagebrush may return to pre-burn density and cover within 15 to 20 years following fire, but establishment after severe fires may proceed more slowly and can take up to 50 years (Bunting et al., 1987; Ziegenhagen, 2003; Miller & Heyerdahl, 2008; Ziegenhagen & Miller, 2009).
The introduction of annual weedy species, like cheatgrass, may cause an increase in fire frequency and eventually lead to an annual-dominated community. Conversely, without fire, sagebrush will increase and the potential for encroachment by juniper also increases. Without fire or changes in management, juniper will dominate the site and mountain big sagebrush will be severely reduced. The herbaceous understory will also be reduced. However, Idaho fescue (Festuca idahoensis) may remain underneath trees on north-facing slopes. The potential for soil erosion increases as the juniper woodland matures and the understory plant community cover declines. Catastrophic wildfire in juniper-controlled sites may lead to an annual weed-dominated site.
Antelope bitterbrush is moderately fire tolerant (McConnell & Smith, 1977). It regenerates from seed and resprouting (Blaisdell & Mueggler, 1956; McArthur et al., 1982). However, sprouting ability is highly variable and is attributed to genetics, plant age, phenology, soil moisture, soil texture, and fire severity (Blaisdell & Mueggler, 1956; Blaisdell et al., 1982; Clark et al., 1982; Cook et al., 1994). Bitterbrush sprouts from a region on the stem approximately 1.5 inches above and below the soil surface. The plant rarely sprouts if the root crown is killed by fire (Blaisdell & Mueggler, 1956). Low-intensity fires may allow bitterbrush to sprout; however, community response also depends on soil moisture levels at time of fire (Murray, 1983). Lower soil moisture allows more charring of the stem below ground level (Blaisdell & Mueggler, 1956), so sprouting will usually be more successful after a spring fire than after a fire in summer or fall (Murray, 1983; Busse et al., 2000; Kerns et al., 2006). If cheatgrass is present, bitterbrush seedling success is much lower. The factor that most limits establishment of bitterbrush seedlings is competition for water resources with the invasive species cheatgrass (Clements & Young, 2002).
Depending on fire severity, rabbitbrush and snowberry (Symphoricarpos spp.) may increase after fire. Rubber rabbitbrush (Ericameria nauseosa) is top-killed by fire, but can resprout after fire and can also establish from seed (Young, 1983). Yellow rabbitbrush (Chrysothamnus viscidiflorus) is top-killed by fire, but sprouts vigorously after fire (Kuntz, 1982; Akinsoji, 1988). Snowberry is also top-killed by fire, but resprouts after fire from rhizomes (Leege & Hickey, 1971; Noste & Bushey, 1987). Snowberry may regenerate well and exceed pre-burn biomass in the third season after a fire (Merrill et al., 1982). If balsamroot or mule-ears is common before fire, it will increase after fire or heavy grazing (Wright, 1985). As cheatgrass increases, fire frequencies will also increase. At frequencies between 0.23 and 0.43 times a year, even sprouting shrubs such as rabbitbrush will not survive (Whisenant, 1990).
The effect of fire on bunchgrasses relates to culm density, culm-leaf morphology, and the size of the plant. The initial condition of bunchgrasses on the site and seasonality and intensity of the fire all factor into the individual species response. For most forbs and grasses, the growing points are located at or below the soil surface. This provides relative protection from disturbances that decrease above ground biomass, such as grazing or fire. Thus, fire mortality is more correlated to duration and intensity of heat, which is related to culm density, culm-leaf morphology, size of plant, and abundance of old growth (Wright, 1971; Young, 1983).
Bluebunch wheatgrass, the dominant grass on these sites, has coarse stems with little leafy material; therefore, the plant’s aboveground biomass burns rapidly and little heat is transferred downward into the crowns (Young, 1983). Bluebunch wheatgrass is typically fairly tolerant of burning, except in May in eastern Oregon (Britton et al., 1990). Uresk et al. (1976) reported burning increased vegetative and reproductive vigor of bluebunch wheatgrass. Thus, bluebunch wheatgrass is experiences slight damage from fire but is more susceptible to fire damage in drought years (Young, 1983). Most authors classify the plant as undamaged by fire (Kuntz, 1982).
Burning can decrease the vegetative and reproductive vigor of Thurber’s needlegrass (Uresk et al., 1976). Fire can cause high mortality, in addition to reducing basal area and yield of Thurber’s needlegrass (Britton et al., 1990). The fine leaves and densely tufted growth form make this grass susceptible to subsurface charring of the crowns (Wright & Klemmedson, 1965). Although timing of fire highly influences the response and mortality of Thurber’s needlegrass, smaller bunch sizes are less likely to be damaged by fire (Wright & Klemmedson, 1965). Thurber’s needlegrass often survives fire and continues growth or regenerates from tillers when conditions are favorable (Koniak, 1985; Britton et al., 1990). Reestablishment on burned sites is relatively slow due to low germination and competitive ability (Koniak, 1985). Cheatgrass is a highly successful competitor with seedlings of this needlegrass and may preclude reestablishment (Evans & Young, 1978).
Basin wildrye is relatively resistant to fire, particularly fire during the dormant season, as plants sprout from surviving root crowns and rhizomes (Zschaechner, 1985). Miller et al. (2013) reported increased total shoot and reproductive shoot densities in the first year following fire, although by year two there was little difference between burned and control treatments.
The grasses likely to invade the sites in this group are cheatgrass and medusahead. These invasive grasses displace desirable perennial grasses, reduce livestock forage, and accumulate large fuel loads that foster frequent fires (Davies & Svejcar, 2008). Invasion by annual grasses can alter the fire cycle by increasing fire size, fire season length, rate of spread, numbers of individual fires, and likelihood of fires spreading into native or managed ecosystems (D’Antonio & Vitousek, 1992; Brooks et al., 2004). While historical fire return intervals are estimated at 15 to 100 years, areas dominated by cheatgrass are estimated to have a fire return interval of 3 to 5 years (Whisenant, 1990). The mechanisms by which invasive annual grasses alter fire regimes likely interact with climate. For example, cheatgrass cover and biomass vary with climate (Chambers et al., 2007) and are promoted by wet and warm conditions during the fall and spring. Invasive annual species can take advantage of high nitrogen availability following fire because of their higher growth rates and increased seedling establishment relative to native perennial grasses (Monaco et al., 2003).
Livestock/Wildlife Grazing Interpretations:
Sheehy and Winward (1981) studied preferences of mule deer and sheep in a controlled experiment: deer showed the most preference for low sagebrush (Artemisia arbuscula), mountain big sagebrush, big sagebrush (formerly foothill big sagebrush; Artemisia tridentata ssp. xericensis), and silver sagebrush (formerly Bolander silver sagebrush; Artemisia cana ssp. bolanderi) and least preference for black sagebrush (Artemisia nova). In a study by Personius et al. (1987), mountain big sagebrush was the most preferred taxon by mule deer. Fecal samples from ungulates in Montana showed that big horn sheep, mule deer, and elk all consumed mountain big sagebrush in small amounts in winter, while cattle showed no sign of sagebrush use (Kasworm et al., 1984).
Antelope bitterbrush is an important shrub species to a variety of animals, such as domestic livestock, antelope, deer, and elk. Bitterbrush is critical browse for mule deer, domestic livestock, antelope, and elk (Wood et al., 1995). Grazing tolerance of antelope bitterbrush depends on site conditions (Garrison, 1953).
Bluebunch wheatgrass is moderately tolerant of grazing and is very sensitive to defoliation during the active growth period (Blaisdell & Pechanec, 1949; Laycock, 1967; Anderson & Scherzinger, 1975; Britton et al., 1990). In a study, herbage and flower stalk production were reduced with clipping at all times during the growing season; clipping was most harmful, however, during the boot stage (Blaisdell & Pechanec, 1949). Tiller production and growth of bluebunch wheatgrass can be greatly reduced when clipping is coupled with drought (Busso & Richards, 1995). Mueggler (1975) estimated that low-vigor bluebunch wheatgrass may need up to eight years rest to recover. Although an important forage species, it is not always the preferred species by livestock and wildlife.
Thurber’s needlegrass is an important forage source for livestock and wildlife in the arid regions of the West (Ganskopp, 1988). The seeds are apparently not injurious, but grazing animals avoid them when the seeds begin to mature. Sheep, however, have been observed grazing the leaves closely, leaving stems untouched (Eckert & Spencer, 1987). Heavy grazing during the growing season has been shown to reduce the basal area of Thurber’s needlegrass (Eckert & Spencer, 1987). This suggests that both seasonality and utilization are important factors in management of this plant. A single defoliation, particularly during the boot stage, can reduce herbage production and root mass thus potentially lowering the competitive ability of this needlegrass (Ganskopp, 1988).
Basin wildrye is valuable forage for livestock (Ganskopp et al., 2007) and wildlife, but is intolerant of heavy, repeated, or spring grazing (Krall et al., 1971). Basin wildrye is used often as a winter feed for livestock and wildlife since it not only provides roughage above the snow but also cover in the early spring months (Majerus, 1992).
Reduced bunchgrass vigor or density provides an opportunity for Sandberg bluegrass and/or cheatgrass and other invasive species to expand onto or occupy interspaces, leading to increased fire frequency and potentially an annual plant community. Sandberg bluegrass increases under grazing pressure (Tisdale & Hironaka, 1981). It is capable of co-existing with cheatgrass. Excessive sheep grazing favors Sandberg bluegrass; however, where cattle are the dominant grazers, cheatgrass often dominates (Daubenmire, 1970). Thus, depending on the season of use, the type of grazing animal, and site conditions, either Sandberg bluegrass or cheatgrass may become the dominant understory species with inappropriate grazing management.
Long-term disturbance response may be influenced by small differences in landscape topography. Concave areas hold more moisture and may retain deep-rooted perennial grasses, whereas convex areas are slightly less resilient and may have more Sandberg bluegrass present.
Inappropriate grazing practices can be tied to the success of medusahead, but eliminating grazing will not eradicate medusahead if it is already present (Wagner et al., 2001). Sheley and Svejcar (2009) reported that even moderate defoliation of bluebunch wheatgrass resulted in increased medusahead density. They suggested that disturbances such as plant defoliation limit soil resource capture, which creates an opportunity for exploitation by medusahead. Avoidance of medusahead by grazing animals allows medusahead populations to expand. This creates seed reserves that can infest adjoining areas and cause changes to the fire regime. Medusahead replaces native vegetation and cheatgrass directly by competition and suppression; it replaces native vegetation indirectly by an increase in fire frequency.
Medusahead litter has a slow decomposition rate because of its high silica content, allowing it to accumulate over time and suppress competing vegetation (Bovey et al., 1961; Davies & Johnson, 2008).
References:
Akinsoji, A. 1988. Postfire vegetation dynamics in a sagebrush steppe in southeastern Idaho, USA. Vegetation 78(3):151-155.
Beckstead, J., and Augspurger, C. K. 2004. An experimental test of resistance to cheatgrass invasion: limiting resources at different life stages. Biological Invasions 6(4):417-432.
Bentz, B., D. Alston, and T. Evans. 2008. Great Basin Insect Outbreaks. In: J. Chambers, N. Devoe, A. Evenden (eds.). Collaborative Management and Research in the Great Basin -- Examining the issues and developing a framework for action Gen. Tech. Rep. RMRS-GTR-204. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fort Collins, CO. Pages 45-48.
Blaisdell, J. P. 1953. Ecological effects of planned burning of sagebrush-grass range on the Upper Snake River Plains. Technical Bulletin No. 1975. Washington, DC: U.S. Department of Agriculture. 39p.
Blaisdell, J. P. and J. F. Pechanec. 1949. Effects of herbage removal at various dates on vigor of bluebunch wheatgrass and arrowleaf balsamroot. Ecology 30(3):298-305.
Blaisdell, J. P. and W. F. Mueggler. 1956. Sprouting of bitterbrush (Purshia tridentata) following burning or top removal. Ecology 37(2):365-370.
Blaisdell, J. P. R. B. Murray, and E. D. McArthur. 1982. Managing Intermountain rangelands-- sagebrush- grass ranges. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. Gen. Tech. Rep. INT-134. 41 p.
Bovey, W. R., D. Le Tourneau, and C. L. Erickson. 1961. The chemical composition of medusahead and downy brome. Weeds 9(2):307-311.
Bradford, J. B., and W. K. Lauenroth. 2006. Controls over invasion of Bromus tectorum: The importance of climate, soil, disturbance and seed availability. Journal of Vegetation Science 17(6):693-704.
Britton, C. M., G. R. McPherson and F. A. Sneva. 1990. Effects of burning and clipping on five bunchgrasses in eastern Oregon. The Great Basin Naturalist 50(2):115-120.
Brooks, M. L., C. M. D'Antonio, D. M. Richardson, J. B. Grace, J. E. Keeley, J. M. Ditomaso, R. J. Hobbs, M. Pellant, and D. Pyke. 2004. Effects of Invasive Alien Plants on Fire Regimes. BioScience 54(7):677-688.
Burkhardt, J. W. and E. W. Tisdale. 1969. Nature and successional status of western juniper vegetation in Idaho. Journal of Range Management 22(4):264-270.
Busse, D., A. Simon, and M. Riegel. 2000. Tree-growth and understory responses to low-severity prescribed burning in thinned Pinus ponderosa forests of central Oregon. Forest Science 46(2):258-268.
Butler, M., R. Simmons, and F. Brummer. 2011. Restoring Central Oregon Rangeland from Ventenata and Medusahead to a Sustainable Bunchgrass Environment – Warm Springs and Ashwood. Central Oregon Agriculture Research and Extension Center. COARC 2010. Pages 77-82.
Clements, C. D. and J. A. Young. 2002. Restoring antelope bitterbrush. Rangelands 24(4):3-6.
Comstock, J. P. and J. R. Ehleringer. 1992. Plant adaptation in the Great Basin and Colorado Plateau. The Great Basin Naturalist 52(3):195-215.
D'Antonio, C. M., and P. M. Vitousek. 1992. Biological invasions by exotic grasses, the grass/fire cycle, and global change. Annual Review of Ecology and Systematics 23:63-87.
Daubenmire, R. 1970. Steppe vegetation of Washington. Technical bulletin 62. Washington Agriculture Experiment Station. 131p.
Davies, K. W., A. M. Nafus, and M. D. Madsen. 2013. Medusahead invasion along unimproved roads, animal trails, and random transects. Western North American Naturalist 73(1):54-59.
Davies, K. W., and D. D. Johnson. 2008. Managing medusahead in the intermountain west is at a critical threshold. Rangelands 30(4):13-15.
Davies, K. W., and T. J. Svejcar. 2008. Comparison of medusahead-invaded and noninvaded Wyoming big sagebrush steppe in southeastern Oregon. Rangeland Ecology and Management 61(6):623-629.
Davies, K. W., C. S. Boyd, D. D. Johnson, A. M. Nafus, and M. D. Madsen. 2015. Success of seeding native compared with introduced perennial vegetation for revegetating medusahead-invaded sagebrush rangeland. Rangeland Ecology & Management 68(3):224-230.
Dobrowolski, J. P., M. M. Caldwell, and J. H. Richards. 1990. Basin hydrology and plant root systems. Pages 243-292 in C. B. Osmond, L. F. Pitelka, and G. M. Hidy (eds.). Plant biology of the basin and range. Springer-Verlag, New York.
Furbush, P. 1953. Control of Medusa-Head on California Ranges. Journal of Forestry 51(2):118-121.
Furniss, M. M. and W. F. Barr. 1975. Insects affecting important native shrubs of the northwestern United States Gen. Tech. Rep. INT-19. Intermountain Forest and Range Experiment Station, U.S. Department of Agriculture, Forest Service. Ogden, UT. 68p.
Harris, G. A. 1967. Some Competitive Relationships between Agropyron spicatum and Bromus tectorum. Ecological Monographs 37(2):89-111.
Hironaka, M. 1994. Medusahead: natural successor to the cheatgrass type in the northern Great Basin. Proceedings of Ecology and Management of Annual Rangelands. USDA Forest Service, Intermountain Research Station. Gen. Tech. Report INT-GTR-313. Pages 89-91.
James, J., K. Davies, R. Sheley, and Z. Aanderud. 2008. Linking nitrogen partitioning and species abundance to invasion resistance in the Great Basin. Oecologia 156(3):637-648.
Johnson, B. G.; Johnson, D. W.; Chambers, J. C.; Blank, B. R. 2011. Fire effects on the mobilization and uptake of nitrogen by cheatgrass (Bromus tectorum L.). Plant and Soil 341(1-2):437-445.
Kerns, B. K., W. G. Thies, and C. G. Niwa. 2006. Season and severity of prescribed burn in ponderosa pine forests: implications for understory native and exotic plants. Ecoscience 13(1):44-55.
Klemmedson, J. O., and J. G. Smith. 1964. Cheatgrass (Bromus Tectorum L.). The Botanical Review 30(2):226-262.
Koniak, S. 1985. Succession in pinyon-juniper woodlands following wildfire in the Great Basin. The Great Basin Naturalist 45:556-566.
Kuntz, D. E. 1982. Plant response following spring burning in an Artemisia tridentata subsp. vaseyana/ Festuca idahoensis habitat type. Moscow, ID: University of Idaho. 73p. Thesis.
Lett, M. S., and A. K. Knapp. 2005. Woody plant encroachment and removal in mesic grassland: Production and composition responses of herbaceous vegetation. American Midland Naturalist 153(2):217-231.
Mack, R. N., and D. Pyke. 1983. The Demography of Bromus Tectorum: Variation in Time and Space. Journal of Ecology 71(1):69-93.
MacMahon, J. A. 1980. Ecosystems over time: succession and other types of change. In: Waring, R., (ed.) Proceedings—Forests: fresh perspectives from ecosystem analyses. Biological Colloquium. Corvallis, OR: Oregon State University. Pages 27-58.
Majerus, M. E. 1992. High-stature grasses for winter grazing. Journal of Soil and Water Conservation 47(3):224-225.
Mangla, S., R. Sheley, and J. J. James. 2011. Field growth comparisons of invasive alien annual and native perennial grasses in monocultures. Journal of Arid Environments 75(2):206-210.
McArthur, E. D., A. Blaner, A. P. Plummer, and R. Stevens. 1979. Characteristics and hybridization of important Intermountain shrubs: 3. Sunflower family. U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. Research Paper INT-177 43.
Miller, H. C., Clausnitzer, D., and Borman, M. M. 1999. Medusahead. In: R. L. Sheley and J. K. Petroff (eds.). Biology and Management of Noxious Rangeland Weeds. Corvallis, OR: Oregon State University Press. Pages 272-281.
Miller, R. F., and Tausch, R. J. 2001. The Role of Fire in Juniper and Pinyon Woodlands: A Descriptive Analysis. In: Galley. K. M. and T. P. Wilson (eds.), Fire Conference 2000: The First National Congress on Fire Ecology, Prevention, and Management; Tallahassee, FL: Tall Timbers Research Station, San Diego, CA, USA. Pages 15-30.
Miller, R. F., J. C. Chambers, D. A. Pyke, F. B. Pierson, and C. J. Williams. 2013. A review of fire effects on vegetation and soils in the Great Basin region: response and ecological site characteristics. Gen. Tech. Rep. RMRS-GTR-308. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 126p.
Monaco, T. A., Charles T. Mackown, Douglas A. Johnson, Thomas A. Jones, Jeanette M. Norton, Jay B. Norton, and Margaret G. Redinbaugh. 2003. Nitrogen effects on seed germination and seedling growth. Journal of Range Management 56(6):646-653.
Murray, R. 1983. Response of antelope bitterbrush to burning and spraying in southeastern Idaho. In: Tiedemann, Arthur R.; Johnson, Kendall L., (comps.). Proceedings: Research and management of bitterbrush and cliffrose in western North America. Ogden, UT: US Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. Gen. Tech. Rep. INT-152. Pages 142-152.
Noste, N. V. and C. L. Bushey. 1987. Fire response of shrubs of dry forest habitat types in Montana and Idaho. Gen. Tech. Rep. INT-239. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 22 p.
Pellant, M., and C. Hall. 1994. Distribution of two exotic grasses in intermountain rangelands: status in 1992. USDA Forest Service Gen. Tech. Rep. INT-GTR-313S. Pages 109-112.
Rice, P. M. 2005. Medusahead (Taeniatherum caput-medusae (L.) Nevski). In: C. L. Duncan and J. K. Clark (eds.). Invasive plants of range and wildlands and their environmental, economic, and societal impacts. Weed Science Society of America, Lawrence, KS.
Sheley, R. L., E. A. Vasquez, A. Chamberlain, and B. S. Smith. 2012. Landscape-scale rehabilitation of medusahead (Taeniatherum caput-medusae)-dominated sagebrush steppe. Invasive Plant Science and Management 5(4):436-442.
Sheley, R. L., and Svejcar T. J. 2009. Response of bluebunch wheatgrass and medusahead to defoliation. Rangeland Ecology & Management 62(3):278-283.
Tisdale, E. W. and M. Hironaka. 1981. The sagebrush-grass region: A review of the ecological literature. Bulletin 33. University of Idaho, Forest, Wildlife and Range Experiment Station. Moscow, ID. 31p.
Vollmer, J. L., and J. G. Vollmer. 2008. Controlling cheatgrass in winter range to restore habitat and endemic fire United States Department of Agriculture, Forest Service. RMRS-P-52. Pages 57-60.
Wagner, J. A., R. E. Delmas, J. A. Young. 2001. 30 years of medusahead: return to fly blown-flat. Rangelands 23(3):6-9.
Whisenant, S. 1999. Repairing Damaged Wildlands: a process-orientated, landscape-scale approach (Vol. 1). Cambridge, UK: Cambridge University Press. 312p.
Wright, H. A. 1985. Effects of fire on grasses and forbs in sagebrush-grass communities. In: K. D. Sanders and J. Durham, (eds.). Rangeland Fire Effects: A Symposium. 1984, November 27-29. USDI-BLM, Boise, ID. Pages 12-21.
Young, R. P. 1983. Fire as a vegetation management tool in rangelands of the Intermountain region. In: Monsen, S.B. and N. Shaw (eds). Managing Intermountain rangelands—improvement of range and wildlife habitats: Proceedings. 1981, September 15-17; Twin Falls, ID; 1982, June 22-24; Elko, NV. Gen. Tech. Rep. INT-157. Ogden, UT. U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. Pages 18-31.
Young-Mathews, A., and S. R. Winslow. 2010. Plant Guide for beardless wildrye (Leymus triticoides). USDA-NRCS, editor. Plants Materials Center, Lockeford, CA.
Ziegenhagen, L. L. 2003. Shrub reestablishment following fire in the mountain big sagebrush (Artemisia tridentata Nutt. ssp. vaseyana (Rydb.) Beetle) alliance. M.S. Oregon State University.
Zschaechner, G. A. 1985. Studying rangeland fire effects: a case study in Nevada. In: K. Sanders and J. Durham, (eds.). Rangeland Fire Effects: A Symposium. 1984, November 27-29. USDI-BLM Idaho State Office, Boise, ID. Pages 66-84.
Major Land Resource Area
MLRA 023X
Malheur High Plateau
Subclasses
- R023XY007NV–LOAMY 14-16 P.Z.
- R023XY015NV–STONY LOAM 12-14 P.Z.
- R023XY016NV–SOUTH SLOPE 12-16 P.Z.
- R023XY018NV–STONY SOUTH SLOPE 12-16 P.Z.
- R023XY022NV–WELL DRAINED FAN 12-14 P.Z.
- R023XY041NV–LOAMY 12-14 P.Z.
- R023XY042NV–GRANITIC SOUTH SLOPE 12-14 P.Z.
- R023XY043NV–GRANITIC SLOPE 14-16 P.Z.
- R023XY094NV–ASHY SLOPE 12-14 P.Z.
- R023XY098NV–DEEP LOAMY 10-12 P.Z.
- R023XY302OR–SOUTH SLOPES 12-16 PZ
- R023XY310OR–NORTH SLOPES 12-16 PZ
- R023XY314OR–GRAVELLY NORTH SLOPES 12-16 PZ
- R023XY316OR–DROUGHTY LOAM 11-13 PZ
- R023XY318OR–LOAMY 12-16 PZ
- R023XY320OR–JUNIPER SOUTH SLOPES 12-16 PZ
- R023XY321OR–DEEP LOAMY 12-16 PZ
- R023XY404OR–DEEP NORTH 12-18 PZ
- R023XY516OR–STONY LOAM 10-12 PZ
- R023XY612OR–PUMICE SOUTH SLOPES 10-14 PZ
- R023XY613OR–PUMICE NORTH SLOPES 10-12 PZ
Correlated Map Unit Components
21500843, 21501125, 21500836, 21501507, 21501149, 21500878, 21501506, 21500868, 21500717, 21501139, 21501109, 21501239, 21500305, 21500310, 21500893, 21500443, 21500450, 21500459, 21500461, 21500466, 21501098, 21500289, 21500601, 21501251, 21500920, 21500697, 21500626, 21500938, 21501389, 21501529, 21501461, 21501521, 21500927, 21500773, 21501012, 21500723, 21500708, 21500970, 21501314, 21501071, 21501566, 21501549, 21500709, 21501307, 21500627, 21501299, 21500676, 21500771, 21501462, 21501459, 21500703, 21500559, 21500564, 21500553, 21500951, 21501075, 21500286, 21501118, 21500682, 21500693, 21500705, 21501180, 21500674, 21500716, 21501541, 21500851, 21500699, 21501553, 21500943, 21500968, 21500960, 21500964, 21500470, 21500566, 21501388, 21500850, 21482028, 21482034, 21482039, 21482043, 21582566, 21582568, 21582564, 21582571, 21589662, 21589674, 21589711, 21590049, 21590098, 21589497, 21589527, 21590127, 21589400, 21589838, 21589635, 21589644, 21589850, 21589861, 21590037, 21589902, 21590042, 21589684, 21589949, 21589873, 21589447, 21589450, 21589457, 21589464, 21589915, 21589589, 21589807, 21589825, 21589766, 21590079, 21589759, 21589675, 21589999, 21589831, 21590128, 21590038, 21589680, 21589879, 21589704, 21589710, 21590067, 21590891, 21590395, 21590597, 21590936, 21590569, 21590571, 21590730, 21590482, 21590307, 21590665, 21590321, 21590480, 21590303, 21590669, 21590315, 21590716, 21590366, 21590870, 21590374, 21590910, 21590940, 21590672, 21590831, 21590334, 21590547, 21590363, 21590915, 21592003, 21591735, 21591732, 21604287, 21604994, 21604294, 21604162, 21604936, 21605198, 21604420, 21604603, 21604177, 21604516, 21604636, 21604408, 21604899, 21604164, 21604172, 21604935, 21605196, 21604637, 21604421, 21729520, 21729021, 21729190, 21729156, 21729422, 21729427, 21729634, 21729425, 21729419, 21729623, 21729626, 21729633, 21729396, 21729402, 21729395, 21729413, 21729400, 21729429, 21728973, 21729142, 21728949, 21730120, 21730121, 21729528, 21729170, 21729019, 21729432, 22176292, 22175048, 22176988, 22175494, 22176912, 22175758, 22176794, 22174973, 22174991, 22175763, 22175447, 22176605, 22176423, 22176939, 22175107, 22175468, 22175469, 22176089, 22176097, 22176819, 22175037, 22175502, 22175546, 22176911, 22175079, 22176951, 22176075, 22176938, 22176960, 22175177, 22176265, 22176842, 22176282, 22176302, 22175284, 22175921, 22177174, 22177180, 22177183, 22177076, 22177348, 22177349, 22177245, 22177477, 22177474, 22177229, 22177126, 22177641, 22168233, 22168250, 22168239, 22168235, 22168283
Stage
Provisional
Contributors
T Stringham (UNR under contract with BLM)
DMP
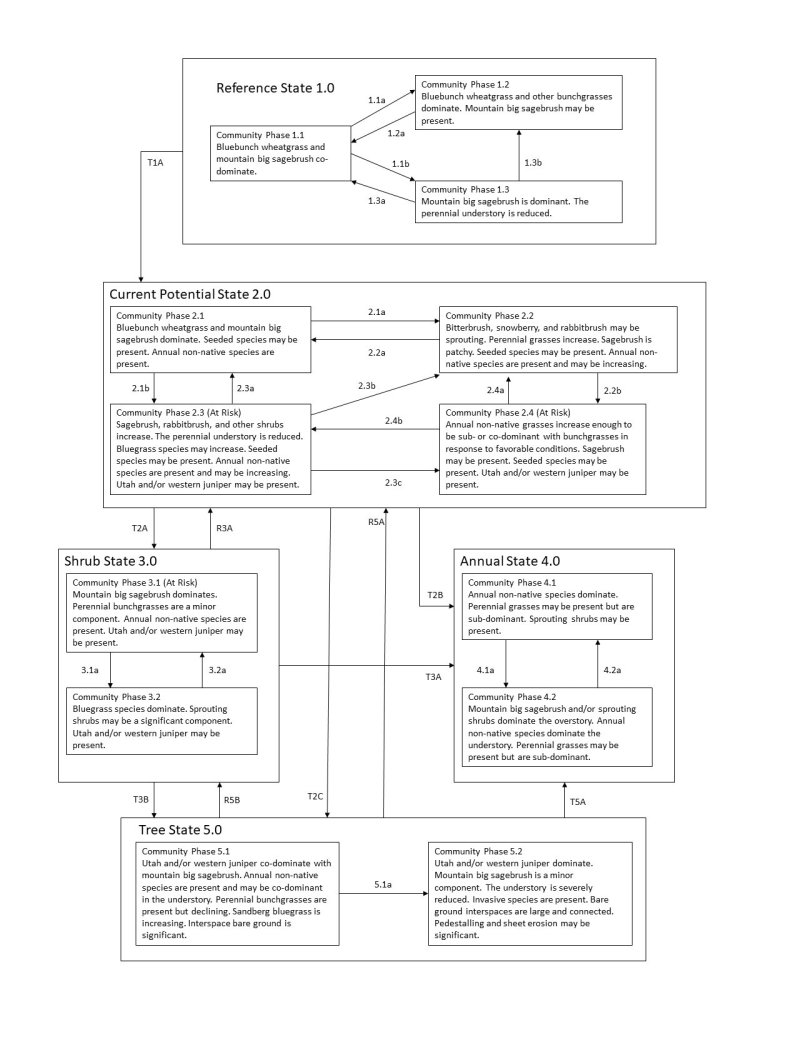

Click on box and path labels to scroll to the respective text.