Ecological site group R023XY912NV
Seasonally Flooded Closed Clay Basins
Last updated: 06/03/2024
Accessed: 04/02/2026
Ecological site group description
Key Characteristics
- Site subject to Ponding or Flooding
- Site is not sodic
- Soils are clayey
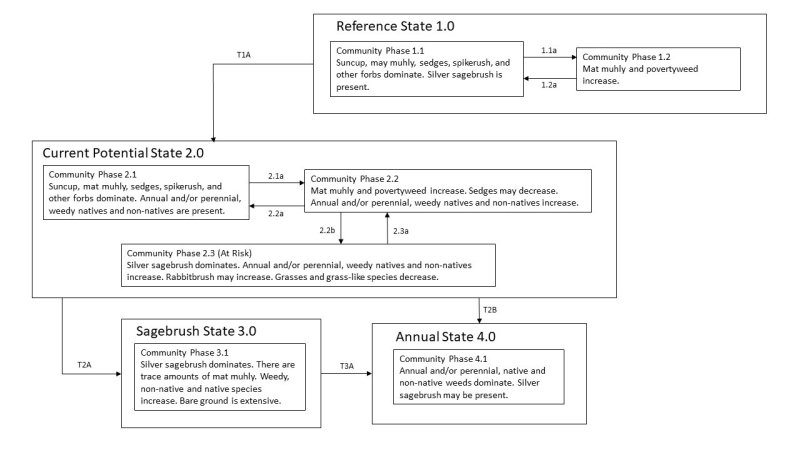
Provisional. A provisional ecological site description has undergone quality control and quality assurance review. It contains a working state and transition model and enough information to identify the ecological site.
Physiography
This group is in enclosed basins at elevations between 5,200 and 6,200 feet. Slopes are less than 5 percent.
These sites are on the lowest settings on the landscape. They receive supplemental water as run in from nearby uplands and are subject to flooding.
Climate
The climate is classified as Cold Semi-Arid in the Koppen Classification System.
The area receives 8 to 14 inches of annual precipitation as snow in the winter and rain in spring and fall. Summers are generally dry.
The frost-free period is 70 to 100 days. The mean annual air temperature is 40 to 46 °F.
Soil features
The soils in the group have clayey textures and do not have root-restrictive layers in the upper 60 inches. Parent materials are alluvium or lacustrine.
Soil temperature regimes are generally frigid with a few mesic map units. Taxonomically, the soils are predominantly Vertisols.
Common soil series in this group include Boulder Lake and Weimer.
Vegetation dynamics
Ecological Dynamics and Disturbance Response:
An ecological site is the product of all the environmental factors responsible for its development. Each site has a set of key characteristics that influence its resilience to disturbance and resistance to invasives. According to Caudle et al. (2013), key characteristics include:
1. Climate factors such as precipitation and temperature.
2. Topographic characteristics such as aspect, slope, elevation, and landform.
3. Hydrologic processes such as infiltration and runoff.
4. Soil characteristics such as depth, texture, structure, and organic matter.
5. Plant communities and their functional groups and productivity.
6. Natural disturbance (fire, herbivory, etc.) regime.
Biotic factors that influence resilience include site productivity, species composition and structure, and population regulation and regeneration (Chambers et al., 2013).
The dominant grass is mat muhly (Muhlenbergia richardsonis): a warm-season, strongly rhizomatous perennial grass that usually grows in loose clumps or mats (USDA, 1988; Penskar & Higman, 1999; Schultz, 2002). Mat muhly reproduces by seed or rhizomes. Mat muhly grows on dry to moist sites and often persists in an area for many years after hydrological modifications lower the water table (USDA, 1988).
Silver sagebrush (Artemisia cana) often grows on deep, poorly drained, often flooded, alluvial soils with high clay contents and seasonally high water tables. Silver sagebrush is an evergreen shrub that often forms colonies from a system of extensive rhizomes (Stubbendieck et al., 1992). The root system of silver sagebrush consists of a taproot with lateral roots and rhizomes, usually located within a few inches of the soil surface. Silver sagebrush sprouts more vigorously than all other sagebrush (Wright et al., 1979). Silver sagebrush can sprout from roots, rhizomes, and the root crown after disturbance (Ellison & Woolfolk, 1937; Whitson et al., 1999; Blaisdell et al., 1982). It readily layers, meaning it can generate adventitious roots from branches that touch the soil (Blaisdell et al., 1982). Silver sagebrush is also capable of reproducing from seeds (Whitson et al., 1999).
Silver sagebrush is a host species for the sagebrush defoliator, Aroga moth (Aroga websteri) (Henry, 1961; Gates, 1964; Hall, 1965). However, it remains unclear whether the moth causes significant damage or mortality to individual plants or entire silver sagebrush stands. Severe drought can kill the crowns of entire stands of silver sagebrush; after drought, however, silver sagebrush can rapidly regrow due to its vigorous sprouting ability (Ellison & Woolfolk, 1937).
Periodic drought regularly influences sagebrush ecosystems. Drought duration and severity have increased throughout the 20th century in much of the Intermountain West. Major shifts away from historical precipitation patterns have the greatest potential to alter ecosystem function and productivity. Species composition and productivity on sites can be altered by the timing of precipitation and water availability within the soil profile (Bates et al., 2006).
The Great Basin sagebrush communities have high spatial and temporal variability in precipitation, both among years and within growing seasons. The Wet Clay Basin ecological site is subject to both periodic drought and flooding that influence the vegetative community from year to year. Many of these sites have been altered since settlement by developments such as dams or dug-out “troughs.” These impoundments and ditches alter the hydrology by changing the area in which water can be captured. If a dug-out lowers the water table, silver sagebrush will increase. If a dam captures more water than a site naturally would, there may be less vegetation on the site due to excessive ponding.
This ecological site group has moderate resilience to disturbance and resistance to invasion. Significant year-to-year variation in ponding and depth to water table are primary drivers for above-ground biomass production. Surface alteration, prolonged drought, or prolonged flooding decrease resilience and increase the probability of annual or perennial weed invasion. Four possible alternative stable states have been identified for this ecological site.
Fire Ecology:
Historically, fire likely was a rare occurrence on these ecological sites. The fire return interval for these ecological sites is a function of the surrounding upland sagebrush sites’ capability to carry fire and prior year rainfall and ponding duration that affect the production of fine fuels. The Wet Clay Basin ecological site in MLRA 23 is often embedded within a larger landscape of Lahontan sagebrush (Artemisia arbuscula ssp. longicaulis). Fire return intervals are largely unknown for Lahontan and low sagebrush but have been estimated at 100 to 200 years in the similar black sagebrush (Artemisia nova) ecosystem (Kitchen & McArthur, 2007). Fires were probably historically patchy due to the low productivity of these sagebrush sites. Sites in this group are unlikely to burn in wet years, and the native vegetation is generally resistant to fire damage.
Mat muhly is resistant to damage from fire because the rhizome buds are insulated by soil (Benedict, 1984). A few studies have observed that fire in the spring stimulates flowering (Anderson & Bailey, 1980; Pemble et al., 1981). However, there is little other documentation of this plant’s fire response.
Beardless wildrye (Leymus triticoides) is a minor component on these sites. It may increase after fire due to its aggressive creeping rhizomes (Monsen et al., 2004). Nevada bluegrass is generally not damaged by wildfire due to its short, tufted growth form and low-density panicles (Monsen et al., 2004). The lack of litter build-up and early dormancy typically preclude extensive damage to the buds, but early fires during dry years may be more damaging (Kearney et al., 1960). Cover of Nevada bluegrass may increase following wildfire (Blackburn et al., 1971). Similarly, Sandberg bluegrass (Poa secunda)—another minor component of these sites—has been found to increase following fire likely due to its low stature and productivity (Daubenmire, 1975). Overall, the grass components of this ecological site group possess structural attributes that suggest high resiliency to fire.
Silver sagebrush is less sensitive to fire than other sagebrush species due to its ability to resprout. Silver sagebrush is capable of resprouting from roots and rhizomes when top growth is destroyed (Cronquist et al., 1994; Blaisdell et al., 1982; Whitson et al., 1999). Silver sagebrush also reproduces by seed. Seedling establishment can occur in the years after fire if the growing season is favorably wet (Wambolt et al., 1989). White and Currie (1983) found spring and fall burning both resulted in complete top-kill of silver sagebrush regardless of fire intensity. However, spring burning when soil moisture was high and before plants began rapid stem growth resulted in low mortality and vigorous sprouting. Fall burning resulted in mortality of 40 to greater than 70 percent of the silver sagebrush plants, suggesting summer wildfires could cause substantial stand death. Post-fire recovery and resilience is primarily influenced by pre-fire site conditions, fire severity, post-fire weather, and land use that relates to vegetation recovery. Sites with low cover of native perennial grasses and forbs typically have reduced resiliency following disturbance and are less resistant to invasion or increases in cheatgrass or other weedy species (Miller et al., 2013).
The dominant forb on these sites, suncup (Camissonia sp.), is unlikely to be affected by fire because it is low-growing and dries up relatively early in the season, before fire is a risk. Povertyweed—a native, perennial, rhizomatous forb—increases following fire due to its prolific seed production and resprouting ability. Povertyweed possesses characteristics of early seral species that are capable of rapidly increasing on disturbed sites (Whitson et al., 1999).
Livestock/Wildlife Grazing Interpretations:
The landscape position of the ecological sites in this group typically provides additional soil moisture for extended plant growth (compared to drier conditions in the surrounding sagebrush landscape), increasing the attractiveness of these areas for animals seeking forage. There is potential for soil damage if grazing occurs when soils are saturated with water, generally in the spring. Mat muhly withstands heavy grazing because of its sod-forming growth form (USDA, 1988). It is a short-statured plant with stems that are typically 3 to 8 inches long and many basal and stem leaves 0.5 to 2 or more inches long (USDA, 1988).
On the drier areas of these ecological sites, bluegrasses and beardless wildrye may be dominant. Nevada bluegrass is very palatable and is preferred by both domestic livestock and wildlife during the spring and early summer. It has reported crude protein levels of over 17 percent (Monson et al., 2004). In today’s botanical climate, Nevada bluegrass and Sandberg bluegrass are no longer differentiated taxonomically. However, these two grasses typically grow in different ecological niches. Nevada bluegrass grows on locations with greater soil moisture during the growing season. Sandberg bluegrass increases under grazing pressure due to its early dormancy and short stature (Tisdale & Hironaka, 1981).
Silver sagebrush can provide an important source of browse and is used by livestock and big game when other food sources are scarce (Kufeld et al., 1973; Wasser, 1982; Cronquist et al., 1994). In fall and winter feeding trials, silver sagebrush was among the most preferred sagebrush species for mule deer and sheep (Sheehy & Winward, 1981). However, silver sagebrush is an aggressive colonizer and can occupy areas at high densities due to its ability to resprout from the crown and spread by rhizomes (Monsen et al., 2004). Therefore, silver sagebrush can increase significantly under inappropriate grazing management on these sites.
Povertyweed is a weedy, native, perennial forb with early seral characteristics such as high seed production that allow it to spread rapidly in disturbed areas (Whitson et al., 1999). Povertyweed can increase when perennial grasses decrease or bare ground increases as a result of excessive mechanical damage to the perennial grasses or soil during wet periods.
In general, inappropriate grazing by domestic livestock or feral horses can cause Nevada bluegrass to decrease and mat muhly to initially increase. Continued deterioration leads to a decrease in mat muhly an increase in povertyweed, silver sagebrush, and annual and perennial weedy forbs.
Hydrologic Modification:
These sites receive additional moisture from runoff from adjacent sites. Off-site and on-site activities can impact hydrology. Years of extreme drought lower the water table. Excessive large animal use during wet periods can cause pugging, root shear, and hummock formation, an increase in bare ground, and modification of infiltration rates. Modifications such as dams, dug-outs, or ditches cause sites to dry, resulting in an increase of weedy annual and perennial forbs and a loss of perennial grasses and potentially silver sagebrush.
References:
Anderson, H. G., and A. W. Bailey. 1980. Effects of annual burning on grassland in the aspen parkland of east-central Alberta. Canadian Journal of Botany 58(8):985-996.
Benedict, N. B. 1984. Classification and dynamics of subalpine meadow ecosystems in the southern Sierra Nevada. California riparian systems: Ecology, conservation, and productive management, edited by R. E. Warner and K. M. Hendrix:92-95.
Blackburn, W. H., R. E. Eckert, Jr., and P. T. Tueller. 1971. Vegetation and soils of the Rock Springs Watershed. University of Nevada, Agricultural Experiment Station, Reno, NV. 116 p.
Blaisdell, J. P., R. B. Murray, and E. D. McArthur. 1982. Managing intermountain rangelands-sagebrush-grass ranges. Gen. Tech. Rep. INT-134. U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station, Ogden, UT. 41 p.
Cronquist, A., A. H. Holmgren, N. H. Holmgren, J. L. Reveal, and P. K. Holmgren. 1994. Intermountain Flora Vascular Plants of the Intermountain West, U.S.A. The New York Botanical Garden, Bronx, New York.
Daubenmire, R. 1975. Plant succession on abandoned fields, and fire influences in a steppe area in southeastern Washington. Northwest Science 49(1):36-48.
Hall, R. C. 1965. Sagebrush defoliator outbreak in Northern California. Res. Note PSW-RN-075. U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station, Berkeley, CA. 12 p.
Henry, J. E. 1961. Biology of the sagebrush defoliator Aroga websteri Clarke in Idaho. M.S. Thesis. University of Idaho, Moscow, Idaho.
Kearney, T. H., R. H. Peebles, and collaborators. 1960. Arizona Flora. University of California, Berkeley, CA.
Kitchen, S. G. and E. D. McArthur. 2007. Big and black sagebrush landscapes. In: S. Hood, M. Miller (eds.). Fire ecology and mangement of the major ecosystems of southern Utah. Gen. Tech. Rep. RMRS-GTR-202. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fort Collins, CO. Pages 73-95.
Miller, R. F., J. C. Chambers, D. A. Pyke, F. B. Pierson, and C. J. Williams. 2013. A Review of Fire Effects on Vegetation and Soils in the Great Basin Region: Response and Ecological Site Characteristics. Gen. Tech. Rep. RMRS-GTR-308. Fort Collins CO: U.S. Department of Agriculture, United States Forest Service, Rocky Mountain Research Station, Fort Collins, CO. 126 p.
Monsen, S. B., R. Stevens, and N. L. Shaw. 2004. Grasses. Pages 295-424 in Restoring western ranges and wildlands, vol. 2. Gen. Tech. Rep. RMRS-GTR-136. USDA, Forest Service, Rocky Mountain Research Station, Fort Collins, CO.
Pemble, R. H., G. L. Van Amburg, and L. Mattson. 1981. Intraspecific variation in flowering activity following a spring burn on a northwestern Minnesota prairie. In: R. L. Stuckey and K. J. Reese,(eds.). Proceedings of the Sixth North American Prairie Conference. 1978, August 12-17. College of Biological Sciences, Ohio State University, Columbus, OH. Pages 235-240.
Penskar, M. R., and P. J. Higman. 1999. Special plany abstract for Muhlenbergia richardsonis (mat muhly). Page 2 in Michigan Natural Features Inventory (editor). Lansing, MI.
Sampson, A., A. Chase, and D. Hedrick. 1951. California grasslands and range forage grasses. Bulletin 724. California Agricultural Experiment Station. 130 p.
Sampson, A.W., B.S. Jespersen. 1963. California range brushlands and browse plants. Berkeley, CA: University of California, Division of Agricultural Sciences, California Agricultural Experiment Station, Extension Service, Berkeley, CA. 162 p.
Schultz, J. 2002. Conservation Assessment for Mat Muhly (Muhlenbergia richardsonis) (Trin.) Rydb. USDA Forest Service, Eastern Region, Excanaba, MI. 31 p.
Tisdale, E. W. and M. Hironaka. 1981. The sagebrush-grass region: A review of the ecological literature. Bulletin 33. University of Idaho, Forest, Wildlife and Range Experiment Station. Moscow, ID. 31p.
[USDA] US Department of Agriculture, Forest Service. 1988. Range Plant Handbook. Dover Publications, Inc., New York, NY. Reprint. Originally published: Washington D. C. Government Printing Office, 1937. 816 p.
Wambolt, C. L., T. Walton, and R. S. White. 1989. Seed dispersal characteristics of plains silver sagebrush. Prairie Naturalist 21(3):113-118.
Whitson, T. D., L. C. Burrill , S. A. Dewey , D. W. Cudney , B. E. Nelson , R. D. Lee , and R. Parker. 1999. Silver sagebrush Artemisia cana Pursh., Big sagebrush Artemisia tridentata Nutt. Pages 62–63, 68–69. In: T. D. Whitson (ed.), Weeds of the west. Western Society of Weed Science, Newark,CA.
Wright, H. A., L. F. Neuenschwander, and C. M. Britton. 1979. The role and use of fire in sagebrush-grass and pinyon-juniper plant communities: A state-of-the-art review. Gen. Tech. Rep. INT-58. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station. 48 p.
Major Land Resource Area
MLRA 023X
Malheur High Plateau
Correlated Map Unit Components
21659972, 21659295, 21660327, 21660329, 21660335, 21659293, 21660498, 21500314, 21501382, 21501602, 21500578, 21501384, 21500976, 21589942, 21589472, 21589466, 21589392, 21589940, 21589470, 21590590, 21590529, 21604337
Stage
Provisional

Click on box and path labels to scroll to the respective text.